Christian Churches of God
[B9A]
L’Évolution et la Théorie
de l''Origine Africaine de l'Homme'
(Édition 1.0
20110116-20110116)
La théorie de l’Évolution
postule que l’homme moderne est apparu en Afrique et s’est répandu à travers
le monde à partir de ce continent il y a 78000 ans. Le récit biblique, quant
à lui, décrit un processus différent. Ces deux affirmations ne peuvent pas
être toutes deux exactes.
Christian Churches of God
Courriel :
secretary@ccg.org
(Copyright
ã
2011 Wade Cox)
(Tr. 2011,
2026, rév. 2026)
Ce document peut être librement copié et distribué, à condition d’être
reproduit intégralement, sans aucune modification ni suppression. Le nom
et l'adresse de l'éditeur ainsi que la mention de copyright doivent être
inclus. Aucun frais ne peut être imposé aux destinataires des copies
distribuées. De brèves citations peuvent être insérées dans des articles
et des revues critiques sans violation du copyright.
Ce document est disponible
sur les pages du World Wide Web :
http://logon.org/ et
http://ccg.org/
L’Évolution et la Théorie de l'« Origine Africaine
de l'Homme » [B9A]
Le
présent exposé se propose d’examiner
les manœuvres des généticiens
Évolutionnistes concernant la conjecture dite l'« Origine Africaine de
l'Homme » (ou « Out of Africa ») et les prétentions qui visent à faire
remonter l'origine de l'homme jusqu’aux Cro-Magnons, puis à une période
située il y a 78.000 ans avant le présent (AP) en Afrique. Pour ce faire,
ces chercheurs ont arbitrairement fusionné les lignées de Cham avec les deux
autres lignées de Sem et de Japhet, en prétendant que tant Sem que Japhet
sont descendus tous deux des lignées de Cham. Cette approche a pour effet de
regrouper les fils de Noé et d’utiliser Cham pour étendre la lignée, par le
recours à un arbre des Haplogroupes totalement erroné, ils parviennent alors
ainsi à faire remonter cette lignée jusqu’en Afrique et à étirer les dates
jusqu’à 78000 AP, ce qui est rigoureusement inexact. Cette manipulation a
été mise en lumière et signalée et peut être observée sur des sites tels que
www.eupedia.com.
Nous nous
proposons ici de reconstruire cette argumentation afin de démontrer, par
l’examen des séquences DYS correctes, en quoi ce raisonnement est spécieux
et que l’échelle chronologique bien plus courte que celle alléguée, et ce
même en adoptant leur propre séquence erronée des taux de mutation.
L’argument est présenté comme suit. L'ancêtre Y-ADN
originel est désigné comme :
Y, prétendument apparu en Afrique il y a 78000 AP
La première mutation A n'est pas répertoriée dans les
listes européennes. En effet, c'est parce que l’Hg. A n'a aucun lien que ce
soit avec les autres et, avec l’Hg. B, est exclusif à l’Afrique et aux
Afro-Américains.
Voir l’Annexe A
En examinant l'arbre phylogénétique du
chromosome Y-ADN ci-dessous, on constate le scindement de l'arbre dans les
lignées qui interdisent absolument qu’un tel arbre conjectural soit formé.
Il convient tout d'abord de partir de
la racine ancestrale de Noé et de la structure de base des principales
branches des Haplogroupes de Cham. L'haplogroupe (Hg.) A était le premier à
subir une mutation à partir de la lignée ancestrale principale. Hg. A a subi
une mutation aux marqueurs M91 et P97?. Ne conservant aucun des liens de
base retrouvés chez les autres lignées.
La lignée ancestrale principale
s’étend de Noé à Japhet, Cham et Sem via les marqueurs SRY0831, 1 M41 2,
M139, M168, P9.
Les autres lignées de Cham ont subi
une mutation vers les lignées M60, M181, et P85? pour former l’Hg B. Les
haplogroupes majeurs A et B sont plus ou moins confinés à l’Afrique et aux
populations afro-américaines.
Les autres
lignées de Cham, issues des lignées Noahides Centrales (marqueurs M168 et
P9) ont formé l’Hg. C (marqueur RPS4Y M216) pour les lignées de Cush, et en
Hg. DE (marqueurs M145 M213), qui constitue la division YAP couvrant les Hg.
D (Tibétains) et E (lignées Cananéennes et nord-africaines).
Il apparaît donc
manifeste que les principales lignées africaines A et B n'ont strictement
aucune parenté avec les autres lignées non-africaines, si ce n’est le fait
que Hg. E a migré et s’est répandu en Afrique du Nord et au Moyen-Orient,
puis de là en Europe.
Toutes les
lignées dans les descendants de Sem et de Japhet se sont répandues partout à
travers le monde via la lignée issue de l’Hg. F et des mutations P14, M89 et
M213 (selon le Diagramme de Phylogénétique FTDNA de 2005). L'examen révèle
que M213 était commun aux groupes DE ainsi qu’à F etc., mais absent de l’Hg.
C. On peut donc raisonnablement penser que M213 a subi une mutation en M216
dans le groupe C avec l’insertion de RPS4Y, tandis que dans le Hg. DE, M145
a été inséré avant M213, et dans l’Hg. F et des lignées subséquentes, P14 et
M89 ont été insérées avant M213. Ainsi, il pourrait être affirmé que M213
constituait la section finale de la lignée originelle Noahide passant par
Sem, Cham et Japhet. Il s’agit assurément de la lignée correcte de l'arbre
Y-ADN tel que mesuré par la science, quelle que soit la durée – longue ou
courte – qu'elle prétend nécessaire à ces mutations.
Il est ainsi
totalement impossible que les Hg. C, D, E et F ainsi que tous les groupes
subséquents G, H, IJ et K, proviennent de A ou B, pas plus qu’il n’est
possible que D, E, F et les groupes subséquents proviennent de C, comme le
prétendent ces arguments l'« Origine Africaine de l'Homme » (ou « Out of
Africa »).
Les données
historiques bibliques et la nature des mutations des groupes indiquent que
les principales mutations sont survenues en cinq mutations spontanées
distinctes à partir des lignées Noahides principales. Elles ont ensuite
évolué et pris fin en des mutations subséquentes formant d'autres groupes
tribaux/nationaux. Prétendre le contraire relève manifestement d’une fraude
délibérée.
Il est manifeste,
d'après leurs propres tableaux, que les lignées et chronologies sont bidons,
et que tous les événements antérieurs à 42000 AP figurant sur leurs
graphiques le sont également.
La répartition
proposée des autres mutations est tout aussi une conjecture complète et
erronée. Les événements ne peuvent donc pas être fondés même sur la date de
42000 AP.
La répartition des
lignées selon les différentes mutations est en réalité tout à fait possible
dans un intervalle allant des années 2200 AEC (Avant l’Ère Courante) jusqu’à
nos jours, si l'on ignore le graphique artificiellement allongé, si l'on
replace le modèle biblique en perspective, et
si l'on discute correctement et prend en compte
comme il se doit les effets modernes observés de la radiation de base ainsi
que les effets de l'ADNmt sur le génome humain.
L'évolution est un mythe religieux qui doit être
dûment exposé et écarté. Cependant, pour maintenir ce mythe religieux, la
science doit être falsifiée et des éléments clefs négligés (omis) ou
dénaturés.
Ce type de
dénaturation est courant. Un cas récent concerne les découvertes relatives à
des humanoïdes en Russie et en Israël, renvoyant à la découverte de 2003 du
soi-disant ‘Hobbit de Flores’.
La découverte d'un os
de doigt en Russie, dans une grotte en Sibérie, a été datée de 30000 AP,
mais comme à l’accoutumée, le fondement de cette datation n'a pas été
discutée publiquement. Son ADN était exceptionnellement bien préservé, selon
les rapports.
Les scientifiques
commentant ces découvertes récentes en Russie ont fait référence au "Hobbit
de Flores” comme datant de plus de 12000 ans, certains suggérant même plus
de 30000 AP pour une espèce humaine ancienne et pré moderne. Ceci en dépit
du fait que des scientifiques ont déjà examiné la découverte de Flores et
ont prouvé, par son Y-ADN, qu'il s'agissait d'un humain moderne dégénéré
ayant régressé en taille et en capacités en raison de son régime alimentaire
sur cette île. Il s’agissait en réalité d’un exemple de mode de vie à éviter
et de la façon dont une société humaine peut perdre ses capacités.
Ces données furent
néanmoins utilisées par des scientifiques qui, soit connaissaient les faits
et les déformèrent délibérément, soit étaient trop paresseux pour allumer
leur télévision, et n'ont certainement jamais étudié les recherches récentes
sur le sujet. À côté des grands prêtres de la Théorie de l'Évolution, les
moines catholiques romains médiévaux ont l'air de progressistes.
Les résultats,
publiés en décembre 2010 dans la revue
Nature, confirmaient que le spécimen provenait d’une jeune fille qui
n'était ni un "humain moderne" ni un Néandertalien. Au contraire, elle a
censément appartenu à une branche distincte, aujourd’hui éteinte, de l'arbre
phylogénétique humain, que les scientifiques ont nommé
Denisoviens, d’après la Grotte de
Denisova en Sibérie du sud où le fossile a été trouvé.
En outre, il a été
rapporté qu'une molaire retrouvée dans la grotte serait celle d’un individu
Denisovien. Cela semble censément différent morphologiquement des dents des
humains modernes et des Néandertaliens, et ressemblerait prétendument
davantage de près à celles d'ancêtres humains beaucoup plus anciens, tels
que l'Homo Erectus.
Basé sur leur
conjecture concernant la structure de l'ADN, les scientifiques continuent
alors d’affirmer que ces découvertes modifient le récit de l'évolution
humaine. Ils continuent alors à affirmer que les découvertes suggèrent que
les Néandertaliens avaient un groupe sœur asiatique qui s'est détaché pour
suivre sa propre voie évolutive avant de s’éteindre. Le fait que les
Néandertaliens possédaient un système ADN complètement différent et
absolument étranger à celui des humains est ignoré, et cette découverte est
alors appliquée à l'hypothèse selon laquelle ces pré-humains avaient une
relation avec la création pré-Adamique des Néanderthaliens etc., alors que
nous savons qu’ils n’en est rien.
La plupart des
scientifiques pensent que les ancêtres des Néandertaliens ont quitté
l'Afrique il y a entre 300000 et 400000 ans pour s'établir en Europe et en
Eurasie. Les scientifiques allèguent ensuite l'affirmation fausse que,
pendant ce temps, les ancêtres directs des humains modernes, les Homo
Sapiens, sont restés et ont évolué en Afrique.
Ils auraient
prétendument quitté l'Afrique il y a 70000 à 80000 ans et auraient coexisté
pendant un temps avec les Néandertaliens, bientôt appelés à s’éteindre.
Les scientifiques
pensent maintenant que les Denisoviens ont divergé des Néandertaliens au
moment de l'exode originel, il y a entre 300000 et 400000 AP, et se sont
répandus vers l'Est.
Les scientifiques
affirment alors, en totale contradiction avec toutes les preuves ADN, que :
“Tout comme les Néandertaliens, ils semblent s'être croisés avec les humains
modernes.” Nous aurions plus de chance de nous reproduire avec un chimpanzé
qu'avec un Néandertalien. Au moins, les chimpanzés possèdent un système à
huit brins, ce qui n'est pas le cas des Néandertaliens.
Les éléments de
preuve seraient prétendument venus inopinément quand des séquences
génétiques Denisoviennes sont apparues dans l'ADN de Mélanésiens modernes,
habitants des îles du Pacifique. Comment cela pourrait-il se produire,
pourrait-on se demander ? La structure de l’Y-ADN Mélanésien est basée sur
la séquence de l’Hg. K, dont nous savons qu’il est un descendant direct de F
et qu’il existe dans la séquence mentionnée plus haut. Quelle partie
mystérieuse de cette séquence de l’Hg. K serait dérivée, en Orient, de
l'Homo Erectus ? Peut-être qu’un des humains Adamiques de l’Hg. K s'est-il
croisé avec une espèce pré-humaine ; bien qu'il n'y ait aucune preuve du
tout pour étayer cette supposition.
La recherche suggère
que le matériel génétique dérivé des Denisoviens constituerait environ 4 à 6
% du code génétique d'au moins certains Mélanésiens. Est-ce exact ? Quels
sont les marqueurs DYS impliqués et quelle est la base de cette affirmation
? Les Mélanésiens étaient-ils des Orang-outans ? Quel ADNmt est impliqué ?
Nous savons pour un fait que l’ADNmt de tous les humains provient de l’ADNmt
Hg. L. Sur quelle base possible peuvent-ils fonder une telle affirmation ?
La raison pour laquelle ils ne divulguent pas la base de ces allégations est
qu'elles sont invariablement fausses une fois exposées à l'antiseptique
efficace que sont l’air frais et la lumière du soleil.
Cette semaine, une
découverte en Israël de huit dents est également présentée comme étant celle
d’un ancêtre moderne des humains. Ils prétendent qu’elle date de 400000 ans.
Le fait est qu'Israël est un site de découvertes de Néandertaliens, toutes
testées et n’ayant absolument aucun lien avec les humains modernes.
À partir des
découvertes en Israël, il a été possible de démontrer qu'ils ont fait cuire
et ont consommé des plantes et des légumes. Des chercheurs américains
auraient trouvé des grains de matière végétale cuite dans leurs dents.
L'étude est la première à confirmer que le régime alimentaire néandertalien
n’était pas limité à la viande et était plus sophistiqué qu’on ne le pensait
auparavant. La recherche a été publiée dans les Proceedings of the
National Academy of Sciences.
Comme la plupart de
ces affirmations, les preuves à ce jour étaient circonstancielles. Ils
étaient définitivement cannibales et les analyses chimiques des os ont
suggéré qu’ils mangeaient peu ou pas de légumes. Cette dépendance perçue à
la viande avait été avancée par certains comme l'une des raisons pour
lesquelles ces humanoïdes se sont éteints, comme les grands animaux tels que
les mammouths qui ont disparu.
Cependant, une
nouvelle analyse, par des scientifiques, à partir de restes néandertaliens à
travers le monde, a trouvé des preuves directes qui contredisent les études
chimiques. Les chercheurs ont trouvé des grains fossilisés de matière
végétale dans leurs dents, dont une partie était cuite.
Bien que les grains
de pollen aient déjà été trouvés auparavant sur des sites néandertaliens et
certains dans des foyers, ce n’est que maintenant qu’il est accepté qu'il
existe des preuves claires que ces peuples consommaient des aliments
végétaux.
“Nous avons déjà
trouvé des grains de pollen sur des sites néandertaliens, mais on n’a pas pu
déterminer s'ils mangeaient des plantes ou dormaient sur elles, ou quoi que
ce soit d’autre.”
"Mais ici, nous avons
un cas où un peu de plante est dans la bouche, donc nous savons que les
Néandertaliens consommaient cette nourriture" (Professeur Alison Brooks
Université de George Washington à BBC).
Les raisons pour
lesquelles les tests ont été faux expliquent le fait pourquoi la plupart des
découvertes sont loin de la vérité. Elles sont fondées sur des hypothèses
provenant des notions préconçues des analystes eux-mêmes.
Selon le Professeur
Brooks, les tests mesuraient les niveaux de protéine, que les chercheurs ont
supposé provenir de la viande.
"Nous avons eu
tendance à supposer que si vous avez une très haute valeur de protéine dans
le régime alimentaire, cela doit venir de la viande. Mais... il est possible
qu'un peu de protéine dans leur régime alimentaire provienne des plantes,"
a-t-elle déclaré.
Cette étude est
considérée pour être la dernière en date à suggérer que, “ loin d’être des
sauvages brutaux, les Néandertaliens nous ressemblaient plus que nous
l’avions précédemment pensé.”
Cette volonté de
trouver un chaînon manquant et de relier les humains pré-Adamiques aux
humains Adamiques est pour maintenir les mythes de la religion de
l'Évolution, et le fait qu'il existe de preuves ADN absolues démontrant sans
l’ombre d’un doute que nous n’avons rien à voir avec les humains
pré-Adamiques ou eux avec nous, est continuellement ignoré et supprimé.
Un scientifique
moderne, avec tout le bagage de la théorie évolutionniste, est aussi
paralysé que ses prédécesseurs dans le système religieux Trinitaire avant
lui. Ne vous y trompez pas : l'Évolution est une religion basée sur le mythe
et la conjecture.
Nous avons traité de
la lignée des nations ci-dessus, et avons montré en quoi il était impossible
que les humains modernes soient sortis d'Afrique il y a 78000 Ans AP.
L'Annexe B ci-dessous montre l'Arbre phylogénétique du chromosome Y-ADN
jusqu’à l’Haplogroupe F.
Nous allons
maintenant réorganiser l’Arbre Y-ADN de façon à montrer plus facilement ou
clairement les lignées Y-ADN de Noé et de ses fils Sem, Cham et Japhet, et
comment les mutations se sont produites à partir de cette lignée, et comment
elles ont subi davantage de mutations parmi les fils de Sem et de Japhet.
Noé était ‘pur dans
ses générations’ (Genèse 6:9). La [version de la Bible] RSV le rend :
“irréprochable dans sa génération” mais il est considéré que c'était sa
structure génétique à laquelle le texte se référait.
La lignée Y-ADN à
partir d’Adam était essentiellement une structure pré M412, M139 qui a formé
le noyau de la structure d’Y-ADN des humains modernes. Noé a transmis à
chacun de ses fils la capacité racine de M168 et P9. Cham a transmis à
chacun de ses fils la capacité de base ou les conditions préalables
génétiques qui ont permis les mutations qui ont abouti aux Hg. A, B, C et DE
[Hg. veut dire Haplogroupe]. La nature des mutations est telle que certaines
conclusions découlent nécessairement de ces mutations, ainsi que de la
distribution des mutations de l’ADNmt qui ont suivi. Nous pouvons tirer des
conclusions étonnantes de la logique de ces mutations et de cette
distribution.
La première déduction
logique que nous sommes requis de faire est celle de déterminer le nombre de
personnes qui étaient présentes sur l'Arche. La Bible dit qu'il y avait huit
personnes. La réponse à cette question soulève des interrogations sérieuses
sur ce à quoi est les textes font référence. Lorsqu’il est fait référence
aux huit personnes, parlons-nous de quatre hommes et de quatre femmes, ce
qui a été la supposition habituelle des étudiants de la Bible au fils des
ans ? Le fait qu'ils soient supposés monogames est également tenu pour
acquis. Un certain nombre d'objections bibliques et scientifiques s’opposent
à cette vue.
La Bible déclare
aussi qu'à la première récolte de vin, qui devait avoir lieu dans un délai
de trois ans suivant le Déluge, Noé s’est enivré. Il fut ensuite outragé.
Compte tenu du fait que Canaan était en vie et suffisamment âgé pour être
considéré comme responsable de ses actes envers son grand-père Noé, il reçut
une malédiction et une peine sévère. Pour être frappé d'une telle peine,
c'est une question de justice et de loi que Canaan ait dû être d'un âge
suffisamment adulte pour être soumis à un tel jugement. Il était le plus
jeune des fils de Cham. Par conséquent, ils doivent tous avoir été sur
l’Arche ensemble. Compte tenu d'autres considérations concernant les
mutations de l’Haplogroupe, il est probable qu'il y avait huit hommes, à
savoir Noé et ses trois fils, et les autres étaient les quatre fils de Cham.
Cela soulève alors des questions concernant les mères. Étaient-ils tous de
la même femme ou étaient-ils des fils de différentes femmes ? Nous verrons à
partir des mutations des gènes et de l'histoire de la Bible qu'ils ont bien
pu avoir des mères différentes, même si elles avaient une structure d’ADNmt
légèrement similaires, mais néanmoins divergentes.
Nous savons
maintenant avec certitude que l’ADN mitochondrial [ou ADNmt] féminin
original était l’Haplogroupe L. Nous savons aussi avec certitude que
l'originel Hg. L est maintenant confiné à l’Afrique ; de là l'origine de
l’hypothèse l'« Origine Africaine de l'Homme » (ou « Out of Africa »). Il y
a peu de doute que la lignée d’Ève était de peau foncée ou, du moins elle
avait la mélanine et la capacité physiologique pour se développer vers la
femme africaine moderne actuelle. Elle était de l’Haplogroupe L, comme
l’était le groupe originel ADNmt. La science de l'ADN considère que les taux
de mutation sont très lents, en fait bien plus lents qu'ils ne le sont en
réalité. En outre, la science a supposé que l’ADNmt n'affecte pas le génome
humain et ne joue aucun rôle dans les mutations génétiques. Ces deux
prémisses sont aujourd’hui prouvées comme étant totalement fausses. La base
de ces changements est discutée dans le document
L’Origine Génétique des
Nations (No. 265).
En effet, c’était ce
document d’étude qui a identifié la nécessité logique pour les Hgs. I et J
d’avoir été l’Haplogroupe hébreu originel IJ qui s’est ensuite scindé en
deux groupes. Dans un premier temps, les scientifiques ont rejeté cette
hypothèse et ils ont récemment trouvé les liens aux marqueurs S2 et S22 de
la structure ; bien qu'ils aient ensuite déclaré que l’Hg. I était
proto-sémite et l'ont antidaté de 30000 ans.
La quantité de
radiation ambiante affecte le taux de mutation de l'ADN humain. Il a été
prouvé que l'ADNmt humain peut muter à un taux allant jusqu'à une mutation
par génération dans des zones à forte radiation ambiante, comme nous l’avons
vu dans les expériences de Kerala mentionnées dans l'étude ci-dessus. En
outre, il a été démontré que l’ADNmt induit des mutations dans l’ensemble du
génome humain. Ainsi, la radiation ambiante fait muter l’ADNmt à un taux
élevé, tout comme l’ADN-Y. Les mutations à l’ADNmt augmentent les taux de
mutation de l'ADN-Y indépendamment de la radiation ambiante, accentuant
ainsi davantage le taux de mutation.
Donc, nous voyons que
quand ils sont combinés, les deux effets sur la mutation doivent logiquement
augmenter le taux selon une séquence de mutation qui s’accélère en raison de
l'exposition croisée à une mutation supplémentaire dans l’ADNmt. Ainsi, le
taux de mutation peut être plusieurs fois supérieurs à celui supposé par la
science pour justifier ses modèles évolutionnistes. La structure entière de
l'argument repose sur des fausses prémisses connues et doit être logiquement
écartée jusqu'à ce qu’elle soit corrigée adéquatement.
Nous pouvons
également prouver, à partir de résultats ADN connus, que les ADN originels
étaient liés et se sont répandus conjointement avec des groupes de base
d’ADNmt connus. Par exemple, les premières découvertes concernant les
Aborigènes d’Australie révélaient huit vagues de trois Haplogroupes de Y-ADN
et trois Haplogroupes d’ADNmt. Ceux-ci étaient Y-ADN Hg. C4 (se mutant en
C4a et C4b, Hg. K et Rx R1basique. Les Haplogroupes ADNmt féminins
correspondent plus ou moins aux groupes masculins précisément. Pour cette
étude nous pouvons voir que les groupes C4 Chamitiques correspondent à l’Hg.
N Basique féminin, qui est la première mutation de L3.
Ainsi, nous
pouvons déduire que L, étant l'origine, a vu la division présente sur
l'arche comme étant des mutations de L et peut-être L1 ou L2 (bien que L2
puisse avoir été une mutation ultérieure de L) et L3 (qui a muté en N puis
M). Les premières divisions de L3 en N Basique ont dû se produire avant que
C4 ne se déplace vers l’Australie. Ces liens doivent correspondre aux
premiers mouvements des Cushites Hg. C hors du Moyen-Orient vers l’Asie et
ensuite vers l’Australie. Nous pouvons ainsi déduire que la mutation de
[l’Haplogroupe] Hg. N basique a dû se produire à partir de L3 aux tout
premiers temps, soit juste avant, soit juste après le cataclysme du Déluge.
Il est possible que [l’Haplogroupe] Hg. N Basique provient d’au moins une
des épouses des fils de Noé et était apparemment sur l'arche. Comme il était
à la base des mutations, il est probable qu'il était présent dans au moins
deux, et probablement quatre, des épouses. Une autre déduction pourrait être
que, comme N Basique n'est pas un facteur majeur en Afrique, les fils de
Cham, qui étaient des Haplogroupes Y-ADN A et B, ne se sont pas mariés dans
cette mutation mais ont plutôt conservé des épouses des mutations de base L1
et L2.
Pour que L2 se
soit formé, L doit avoir été sur l'arche, ou L2 s’était formé avant elle, et
les trois mutations L1, L2 et L3 étaient présentes. Comme il n'y avait pas
d’autres mutations à partir de L, l’haplogroupe L originel doit avoir vu des
mutations à l'avance, et l’haplogroupe L originel avait probablement cessé
d'exister. Ainsi, il devait y avoir les haplogroupes ADNmt L1 (peut-être
L2), L3, et probablement N Basique sur l'arche. Étant donné que l’ADNmt Hg.
M a subi une mutation à partir d’autres Haplogroupes, tout comme Hg. R a
subi une mutation à partir d’Hg. N, il n'est pas nécessaire pour eux d'avoir
été sur l'arche.
Il est évident qu'il
y a une situation d’entonnoir (goulot d'étranglement) avec L et qu'aucune
autre mutation n'a été observée à partir de celle-ci ou de la structure L1
et L2, et que toutes les mutations ultérieures proviennent des haplogroupes
ADNmt N et M, et subséquemment de N en tant que supergroupe R et les groupes
suivants. Cela a donné lieu à l'affirmation selon laquelle la mutation de N
et M etc. a eu lieu hors de l'Afrique. L'explication biblique est que cette
situation s’est produite au Moyen-Orient et que l'Afrique a été donnée aux
fils de Cham en tant que leur héritage et qu’ils se sont déplacés là après
le Déluge et lors de la dispersion. Le fait que les Y-ADN Hgs. C, D et
certains E ont été trouvés bien à l'extérieur de l'Afrique et se soient
répandus dans le monde ne soutient pas logiquement la théorie "Origine
Africaine de l’Homme”.
En fait, cela
soutient l’inverse de la théorie. La plaque africaine comprend en réalité
l'ouest de la faille de la Jordanie jusqu’à la côte du Liban. Nous pourrions
donc conclure que certains d'entre eux étaient “en Afrique.” Cependant, le
territoire de Cush ou Khus (Koush) était au nord de l'Euphrate. Le fait est
que bibliquement les terres du Jardin d'Éden s’étendaient jusqu’à l'Égypte
et au Nil, et à la côte de la Méditerranée. C'est le candidat logique
probable pour la dispersion, et c'est exactement là que la Bible la situe.
Il est probable que
les épouses des haplogroupes Y-ADN A et B étaient L1 et L2, car ces groupes
sont allés en Afrique ensemble et y sont restés. Les épouses des autres
étaient de l’ADNmt L3 et N basique. L3 ne se trouve nulle part ailleurs
qu'en Afrique, sauf dans des migrations ultérieures connues. Ainsi, N
apparaît comme la principale source d’ADNmt à l’extérieur de l'Afrique. M
peut bien avoir été là aussi mais il apparaît que N se soit répandu sur les
autres et a subi une mutation. Il est probable que l’ADNmt Hg. N était le
plus prolifique des ADNmt parmi les fils de Noé. Les deux autres fils de
Cham peuvent bien s'être mariés avec des femmes de l’Hg. N, tout comme Sem
et Japhet. Cela expliquerait plus facilement la distribution et les
mutations ultérieures.
La présence de N
basique avec C4 indique que les filles de l’Haplogroupe ADNmt Hg. N se sont
mariées très tôt avec les Cushites ou y étaient présentes à l'origine. Notez
aussi qu'il n'y avait aucune conservation du M168 P9 dans l'Y-ADN des Hgs. A
et B. Soit ils sont entrés en Afrique avant l'intervention à Babel et ne
l'avaient pas initialement, soit, alternativement, ils l'ont perdu dans les
activités virales/radiations là-bas. L’Hg. A a subi une mutation à partir de
la séquence initiale, du moins c’est ce qu’il semble.
Ce que nous pouvons
constater, c’est que rien de ce que nous connaissons dans la Science de
l'ADN ne contredit la structure biblique et qu’en fait, cela contribue à
expliquer les questions de la Bible plus efficacement. Les éléments qui
entrent en conflit avec la Bible, une fois examinées, montrent que les
conjectures à long terme sont fausses et que les tentatives d'allonger les
chronologies sont basées sur une pseudo-science frauduleuse. Voir le
graphique à l’Annexe C pour les branches des lignées du système Y-ADN
attribuées aux fils de Noé.
q
Annexe
A
Modèle
Évolutionniste de ‟l’Origine Africaine de l’Homme‟
|
Événement / Mutation
allégué(e) |
Chronologie alléguée (années Avant le Présent) |
|
Puis la lignée aurait subi une mutation en BT |
74 000 AP |
|
Ensuite, la séquence serait passée à CT |
70 000 AP |
|
Cette mutation coïnciderait prétendument avec la
Première Migration hors d'Afrique |
|
|
Puis la lignée se serait divisée en deux branches
: |
|
|
DE |
~65 000 AP |
|
L'autre branche CF (les deux provenant de la
séquence Hg. C) serait apparue |
~58 000 AP |
|
DE se serait séparé hors d'Afrique en Hg. D et E,
formant Hg. E à |
~54 000 AP |
|
CF aurait continué à muter en Hg. F |
~48 000 AP |
|
Hg. F se serait ensuite divisé en : |
|
|
Hg. IJK |
~45 000 AP |
|
et Hg. G |
~32 000 AP |
|
(Hg. G aurait subi une mutation pour former G2a il
y a 12 000 AP, voir ci-dessous) |
|
|
Hg. IJK se serait scindé pour former : |
|
|
Hg. K |
~36 000 AP |
|
et Hg. IJ |
~32 000 AP |
|
Hg. IJ aurait subi une mutation pour former J |
~29 000 AP |
|
et I |
~24 000 AP |
|
Hg. I serait apparu dans la région de l'Allemagne,
ce qui est impossible. Nous examinerons plus tard cette fiction. |
|
|
Hg. K aurait muté en NOP |
~36 000 AP |
|
Hg. K aurait aussi subi une mutation pour former
Hg. R hors d'Afrique |
~30 000 AP |
|
Hg. NOP aurait subi une mutation pour former les
Hgs. NO et P |
~29 000 AP |
|
Hg. NO aurait subi une mutation pour former N |
~22 000 AP |
|
P aurait ensuite subi une mutation pour former Hg.
Q |
~24 000 AP |
|
et Q1a |
~20 000 AP |
|
Hg. I aurait formé Pre I1 après 22 000 AP, puis I2 |
~17 000 AP |
|
G2a se serait formé il y a 12 000 AP et G2a3 se
serait formé |
8 000 AP |
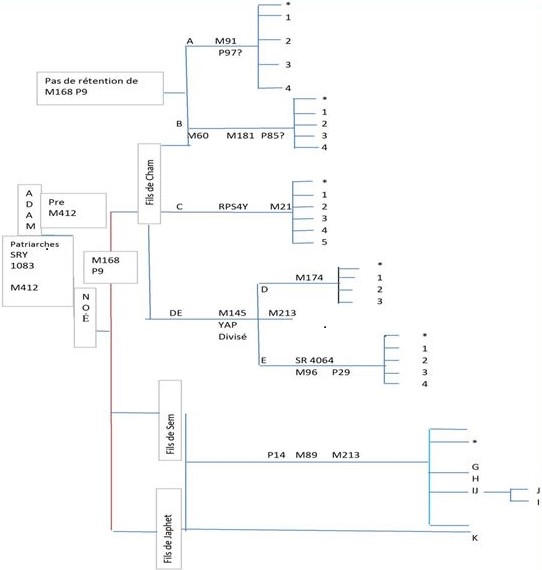
Annexe B
L’Arbre phylogénétique Y-ADN Usuel
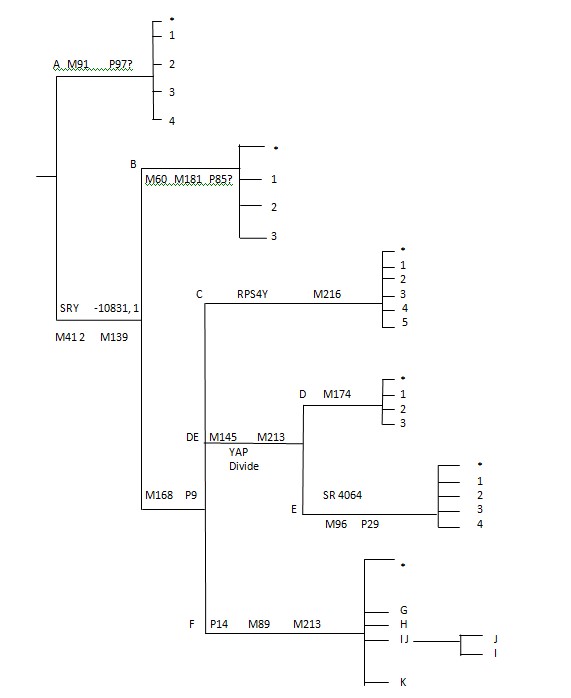
Annexe C
L’Arbre Y-ADN appliqué à la Structure de la Bible