Les Églises Chrétiennes de Dieu
[265]
(Édition
4.5
20060105-20060421-20070804-20071122-20200123)
Christian
Churches of God
PO Box 369, WODEN
ACT 2606, AUSTRALIA
Courriel: secretary@ccg.org
(Copyright © 2006, 2007, 2020 Wade Cox)
(Tr. 2013, rév. 2020)
Cette étude peut être copiée et distribuée
librement à la condition qu'elle le soit en son entier, sans modifications ni
rayures. On doit y inclure le nom, l'adresse de l’éditeur et l'avis des droits
d'auteur. Aucun montant ne peut être exigé des récipiendaires des copies
distribuées. De brèves citations peuvent être insérées dans des articles et des
revues critiques sans contrevenir aux droits d'auteur.
Cette étude est
disponible sur les pages du World Wide Web à:
http://logon.org/ et http://ccg.org/
L’Origine Génétique des Nations [265]
L’un des avancements les plus
importants dans la génétique
humaine est celui de la mesure
de l'ADN des hommes et des
femmes. Les hommes sont mesurés
par le chromosome Y qui est
passé de père en fils en ligne
continue qui subit une mutation
au cours du temps. C'est appelé
l’ADN-Y. Seulement les hommes le
portent par leur chromosome Y,
étant XY. Les femmes ont les
chromosomes XX. La science dit
que ce taux de mutation est
beaucoup plus lent que la Bible
l'exige pour concorder avec son
histoire des nations comme
descendant de Noé.
Le lignage féminin est mesuré en
testant leur ADN Mitochondrial
(appelé ADNmt) que les hommes
autant que les femmes possèdent.
La mitochondrie est regroupée en
Haplogroupes qui sont reliés aux
lignages des femmes dans
lesquels ils surviennent et ce
lignage est passé de la mère à
sa progéniture dans le
chromosome X, que tant l’homme
que la femme possèdent.
L’ADN-Y et l’ADNmt sont mesurés
de deux façons différentes.
L’ADN-Y est mesuré dans ce qui
est appelé les polymorphes. On
attribue à ces polymorphes une
valeur numérique et, selon la
valeur au moment du test, les
sous-groupes qui sont formés
sont appelés
clades et
sous-clades du groupement
global qui est appelé un
Haplogroupe. Ces valeurs
permettent de constater le
changement dans les mutations et
les lignages de l’ADN-Y.
L’ADNmt est déterminé en
dénotant le site polymorphe
comme par exemple 311C,
signifiant qu’une mutation s’est
produite à la paire de base
16,311 et la base qui a changé
ici a été en réalité changée en
cytosine.
Le nombre 16,000 est le point de
commencement pour la
numérotation de l'ADN et ainsi
le 16,000 est annulé et les
nombres utilisés sont les
nombres supérieurs. Donc 16,311
devient 311 et la lettre indique
le produit chimique à ce
moment-là dans le polymorphe.
C'est ce changement du site
polymorphe qui détermine la
généalogie génétique, comme le
parent passe à sa progéniture
les polymorphismes de l'ADN
qu'ils ont avec les mêmes
valeurs numériques ou
semblables. Lorsque testées, ces
valeurs, qui ne sont pas
exactement les mêmes que le
parent, sont désignées comme
mutations. Les valeurs
varient donc et ont déterminé
les groupements tribaux des
nations du monde.
Le système de l’ADN-Y qui a été
attribué au mâle de l'espèce
humaine est groupé dans une
série d’Haplogroupes de A à R.
La mesure étendue habituelle
(utilisant le système Arizona)
est d'habitude de 37 sites en
tant que marqueurs. La mesure de
base est faite pour les douze
premiers, ensuite à 25 et
ensuite jusqu’à 37 de ces sites
polymorphes, ou emplacements
pour déterminer le degré de
relation et l’association des
Haplogroupes. Il pourrait y
avoir quelques 100 marqueurs ou
plus évalués pour des
changements (aussi connus comme
polymorphismes). Un exemple de
ce que pourrait être une
structure de l'ADN-Y typique est
montré ci-dessous :
|
1 |
393 |
x |
|
2 |
390 |
x |
|
3 |
19 * |
x |
|
4 |
391 |
x |
|
5 |
385a |
x |
|
6 |
385b |
x |
|
7 |
426 |
x |
|
8 |
388 |
x |
|
9 |
439 |
x |
|
10 |
389-1 |
x |
|
11 |
392 |
x |
|
12 |
389-2 |
x |
|
13 |
458 |
x |
|
14 |
459a |
x |
|
15 |
459b |
x |
|
16 |
455 |
x |
|
17 |
454 |
x |
|
18 |
447 |
x |
|
19 |
437 |
x |
|
20 |
448 |
x |
|
21 |
449 |
x |
|
22 |
464a ** |
x |
|
23 |
464b ** |
x |
|
24 |
464c ** |
x |
|
25 |
464d ** |
x |
|
26 |
460 |
x |
|
27 |
GATA H4 |
x |
|
28 |
YCA II a |
x |
|
29 |
YCA II b |
x |
|
30 |
456 |
x |
|
31 |
607 |
x |
|
32 |
576 |
x |
|
33 |
570 |
x |
|
34 |
CDY a |
x |
|
35 |
B CDY |
x |
|
36 |
442 |
x |
|
37 |
438 |
x |
*
Aussi connu comme DYS 394
**
Le 19/05/2003, ces valeurs ont
été rajustées à la baisse de 1
point en raison d'un changement
dans la nomenclature de
Laboratoire.
Il y a des valeurs qui, lorsque
testées, sont placées dans la
colonne de droite de la table.
Ces valeurs aident à déterminer
si une personne est tombée dans
un Haplogroupe celte typique ou
un qui se trouve parmi un type
racial sémitique, ou amérindien,
ou polynésien, ou chinois ou
japonais. Pour correctement
déterminer le type, on exécute
ce qui est connu comme un test
SNP (polymorphisme de nucléotide
simple), mais les valeurs des
tests des marqueurs 12, 25 et 37
nous pointent dans la bonne
direction. Aucun groupe national
n'est d’un Haplogroupe génétique
pur à l'exception de quelques
tribus amérindiennes du groupe Q
en Amérique du Sud (à partir des
diagrammes produits par le Dr.
J. D. McDonald). La table
ci-dessus, selon les valeurs
testées, pourrait être celle du
Celte R1b typique de l'Europe ou
des Amérindiens, dont l'ADN
appartient aux deux derniers
Haplogroupes sur l'échelle.
L’ADNmt
La mitochondrie, initialement
mise en séquence en 1981, est
devenue connue sous le nom de la
Séquence de Référence de
Cambridge (CRS). Le CRS a été
utilisé comme une base de
comparaison avec l’ADNmt
individuel. Autrement dit,
n'importe quelle place dans l’ADNmt
individuel qui a une différence
avec le CRS est considérée comme
une mutation. Si un résultat ne
montre aucune mutation du tout,
cela signifie que l’ADNmt
correspond au CRS. Une mutation
se produit : a) lorsqu’une base
remplace une autre base - par
exemple un C (Cytosine) remplace
un A (Adénine) ; b) lorsqu’une
base n'est plus dans cette
position, ou une délétion ; et
c) lorsqu’une nouvelle base est
insérée entre les autres bases
sans en remplacer une autre (une
insertion).
C'est ainsi que l’ADNmt subit
une mutation. L'ADN féminin
original de l'espèce humaine a
été restreint à sept femmes,
mais en Europe seulement. Dans
le monde entier, il y a environ
26 sous-embranchements de
l'arbre du genre humain pour les
lignages des femmes. Donc, on
pourrait débattre que les
lignages des femmes seraient
explicables en termes bibliques
en Europe seulement. L’ADNmt
exige donc l’harmonisation avec
l’histoire Biblique en
expliquant la totalité des 26
lignes féminines.
Nous sommes dépendants de la Science dans son organisation des Haplogroupes. À mesure que les Polymorphes mutent, ils changent les valeurs et donc le taux de mutation peut bien causer un peu de mouvement entre les Haplogroupes pas encore identifiés.
Arbre Simplifié des Haplogroupes
du Chromosome Y
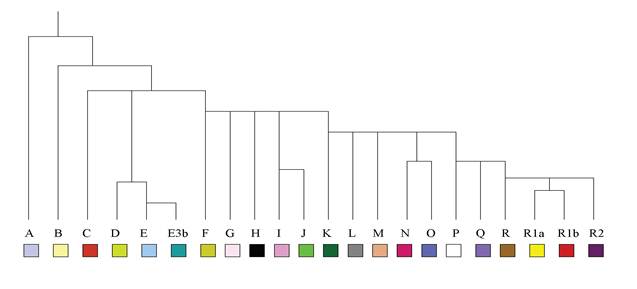
Harmonisation de la Bible et de
la Génétique
Si la Bible doit répondre au
défi posé par cette très
importante découverte génétique,
elle doit alors superposer cette
structure au récit biblique et
expliquer la distribution des
nations dans des termes qui
peuvent tenir compte et répondre
correctement aux défis posés par
les arbres génétiques autant de
l’ADN-Y que de l’ADNmt de
l'espèce.
Les lignages des femmes peuvent
bien avoir un effet sur l’homme
en causant des mutations que
l'on ne connaît pas encore et un
examen de ce système révélera
peut-être aussi quelques aspects
importants. Cependant, à
présent, autre qu’une
recombinaison connue, aucune
conséquence sur les mutations
masculines causées par la femme
n’est scientifiquement
démontrée.
Cependant, nous savons
maintenant que les mitochondries
sont directement responsables
des mutations de la structure
génétique des humains. Elles
s’insèrent dans les gènes actifs
et causent la mutation. Cela est
particulièrement vrai dans l'ADN
qui est endommagé par la
pollution ou la radiation, ce
qui les rend sensibles aux
dommages supplémentaires de
l'ADN mitochondrial parasite.
Les gènes de travail sont plus
vulnérables aux dommages alors
que leur ADN est régulièrement
décompressé à multiplier des
protéines (New Scientist,
18 Septembre 2004, art.
"The enemy within that targets
genes",
p. 11).
"Miria Richetti à l'Institut
Pasteur de Paris et son équipe
ont identifié 211 insertions
d'ADN de mitochondries dans le
génome, 23 d'entre elles sont
nouvelles, 80 pour cent étaient
dans les gènes, mais les gènes
ne représentent que 3 pour cent
du génome. Les cibles sont des
gènes transcrits largement,
probablement parce qu'ils ont
plus de doubles cassures, dit
Richetti" (ibid.).
Jef Boeke à l'Université John
Hopkins pense que l'ADN
Mitochondrial est peu
susceptible d'être une source
majeure de mutation du gène
humain. Cependant, il semblait y
avoir un cas de cela directement
lié à la catastrophe de
radiation de Tchernobyl.
En comparant les séquences de 21
bénévoles de diverses origines
ethniques, l'équipe de Richetti
a trouvé six insertions qui
n'ont pas été partagées par
l'ensemble d'entre eux. (PLoS
Biology, vol. 2, e273). Les
scientifiques ont jugé que
"celles-ci doivent avoir eu lieu
après que nos ancêtres se soient
divisés des ancêtres des
chimpanzés et pourraient donc
donner aux scientifiques des
indices sur les origines des
groupes ethniques et des
migrations historiques"
(ibid.).
Nous sommes convenus que ces
mitochondries sont capables de
muter la structure génétique de
l'homme, et l'incidence des
mariages mixtes avec de
nouvelles mitochondries est
tenue pour provoquer des
mutations à un rythme accéléré.
C'est pourquoi les sociétés
isolées révèlent la structure de
base et moins de changement de
groupes que ne le font les
sociétés plus ouvertes
entremêlées avec d'autres
Haplogroupes mitochondriaux. Par
exemple, les RxR1 parmi les
Aborigènes d'Australie sont
également moins affectés. Ce
n'est pas un indicateur de
temps, mais plutôt un indicateur
de mariages mixtes. La majorité
de l’Haplogroupe C Aborigène,
cependant, est en fait C4 et est
dérivé des groupes de base C qui
se sont propagés le long de la
côte d'Asie du Sud de l'Inde au
Vietnam. Il y a une certaine
base C toujours là dans les
poches, mais les Aborigènes se
sont développés dans le groupe
C4, puis se sont subdivisés en
deux sous-groupes C4 en
Australie. Les Maoris se sont
développés dans le groupe C2 et
les Mongols dans le C3, qui est
également entré dans les
Amériques. Tant les langues
autochtones que maories
contiennent beaucoup de mots
d'emprunt du peuple malais et
indiquent qu'elles ont autrefois
partagé un espace commun et un
groupe linguistique. Les Malais
sont toutefois principalement Hg
O, mais ont quelques autres
groupes. Les Maoris ont quitté
leur île natale ca. 1000 EC,
après la montée des eaux et sont
allés à la Nouvelle-Zélande
pendant la Période Chaude
Médiévale lorsque les mers
étaient plus élevées.
Il semble y avoir un problème
avec ce qui est appelé l'Arbre
Phylogénétique du Chromosome Y
en ce que le
Haplogroupe A est la
première branche manifestement
divergente dans l'arbre et donc
la première à diverger des Hgs B
à R. Tous les Haplogroupes ont
une branche où ils divergent.
Nous regarderons maintenant une
vue d'ensemble du récit
biblique. La Bible dit
simplement que Noé était
parfait dans ses générations
et qu'il a engendré Japhet et
ensuite Cham et ensuite Sem. La
progéniture des fils de Noé est
inscrite et les tribus et la
distribution sont notées et
ainsi nous accordent quelques
indices précieux pour les
comparer au registre de l'ADN.
Les récits Bibliques ont été
interprétés comme ne permettant
seulement que huit personnes sur
l'Arche. Cependant, le récit
peut se référer à huit hommes,
vu que les femmes n'ont jamais
été mentionnées dans les
généalogies antiques sauf dans
des cas très rares et parfois
seulement dans la rature d'un
préfixe du nom des pères
(consulter l’étude
La Généalogie du Messie (No. 119)).
Ce scénario de huit hommes est
probable vu que Canaan était
assez vieux dans la première
moisson de vin après le Déluge
pour avoir sodomisé ou châtré
Noé quand il était ivre de
nouveau vin. Les traditions
juives abordent l'un ou l'autre
des scénarios. C'est la raison
de la malédiction biblique de
Canaan. Certains soutiennent que
c’était Cham, et Canaan a porté
la malédiction plutôt que Cham,
mais cela semble être pour
confiner les huit bouches aux
hommes et aux femmes. Le dernier
cas est important pour le
registre de l'ADN en ce que
seules les femmes des fils
auraient alors continué la
reproduction et l’ADNmt de la
femme de Noé se serait arrêté à
moins qu'il n'y ait eu des
filles de la femme de Noé
présentes aussi, ce qui est
possible si seulement les hommes
sont mentionnés.
Les lignages des hommes étaient
donc Japhet et Sem avec leurs
femmes et Cham et les (quatre)
fils de Cham et leurs femmes
totalisant sept hommes et sept
femmes en plus de Noé et sa
femme.
Cela permettrait alors aussi
huit femmes incluant la femme de
Noé, fournissant ainsi les sept
lignages féminins principaux d’ADNmt,
et cela semble s'accorder avec
les découvertes actuelles des
Haplogroupes d’ADNmt, mais pour
l'Europe seulement. On pourrait
argumenter que quelques lignages
d’ADNmt sont des divisions
postérieures. La preuve
scientifique indique qu'il y
avait sept prétendues "Ève" dans
le bassin génétique de l’ADNmt
pour le Caucasien mais il y a 26
lignes féminines en tout. Donc,
l'explication doit rendre compte
de la séquence complète.
L'argumentation pourrait ainsi
être avancée qu'il y avait 26
femmes sur l'Arche qui ont
continué la reproduction.
Cette opinion placerait la
Science en harmonie directe avec
le récit biblique en ce qui
concerne le lignage des femmes.
Nous constaterons aussi que
lorsque nous regardons le
registre d’ADNmt pour le lignage
des femmes, nous sommes alors
capables d’établir la
correspondance des divisions
féminines avec les divisions
masculines d’ADN-Y. Ainsi nous
pouvons voir la dispersion des
familles de Noé dans leur
distribution dans le monde. Nous
examinerons cet aspect plus
tard.
Le patrimoine génétique de Noé
Noé était reconnu comme étant
pur dans ses générations. La
Bible maintient aussi que les
personnes dans l'Arche étaient
toutes de la famille de Noé.
Ainsi, pour rendre compte
correctement de la diversité
génétique, Noé doit avoir
maintenu la capacité d’avoir une
progéniture génétiquement
distincte et cette progéniture
avait les caractéristiques de la
ligne dont elle est issue mais
pas la séquence complète que Noé
avait à l'origine.
Les opinions conventionnelles
sur les systèmes d’ADN-Y sont
telles qu'elles comptabilisent
35,000 ans dans le passé et
certains disent 60,000 à 130,000
ans et que les niveaux de
mutation sont lents. Cette
opinion étendue est basée sur
des modèles mathématiques, mais
apparemment pour promouvoir un
modèle évolutionniste. En outre,
aucune connaissance n'a tenue
compte de l'impact des divers
systèmes d'ADN féminins. L’ADNmt
est mesuré mais les effets du
chromosome X féminin sur la
structure de l’ADN-Y masculine
sont considérés comme importants
et on argumentera qu'il cause
des mutations dans la structure
de l’ADN-Y à un taux beaucoup
plus rapide que ce que l’on
croit permis par le modèle
actuel.
L’une des histoires connues que
nous pouvons examiner est celle
des Juifs et nous pouvons
identifier un certain nombre de
nations qui viennent d'eux et
dans leur structure d'ADN, nous
pouvons voir beaucoup de peuples
qui ne sont pas réellement des
Sémites selon la connaissance
actuelle. Par exemple, 52 % de
tous les Lévites Ashkénazes ne
sont pas des Sémites selon les
modèles d’ADN-Y et les
explications actuelles. Donc,
nous pourrions avoir la preuve
ici de l’existence d'un grand
nombre de gens convertis au
Judaïsme se mariant dans les
tribus. Selon toute probabilité,
cela pointe à la conversion des
Khazar comme étant la source des
Lévites Ashkénazes. Nous
examinerons cet aspect
ci-dessous.
En outre, nous savons que le
clan Buba du peuple Lemba au
Zimbabwe dans le lignage d’ADN-Y
sont tous Cohen de Lévi et les
Lemba sont principalement de
l’ADN-Y juif, bien qu'ils soient
noirs ayant pris des femmes
africaines après qu'ils ont été
isolés à ou après la captivité
babylonienne. Leur Judaïsme est
beaucoup plus fondamental que le
Judaïsme de post-captivité et
leur détermination du calendrier
est basée sur la conjonction
déterminée à l’avance. Ils ont
maintenu leurs croyances sous
une forme très simple pendant
2,500 ans séparément de Juda.
Toutes les zones de distribution
d'ADN dans cet ouvrage sont en
fonction des populations
indigènes connues avant la
dispersion européenne ca. 1500,
selon les tests.
Pour que Noé puisse être le père
de la structure humaine, il est
reconnu pour avoir eu la
capacité pour la sous-structure
d’ADN-Y suivante vu que tous les
humains sont descendus de lui.
N'importe quel homme sur la
planète aura seulement les
mutations qui signifient
seulement sa branche et son
tracé.
M91, SRY 10831a, M42, M94, M139,
M60, M181, RPS4Y711,
M216, YAP M145, M203, M174, SRY
4064, M96, P29, P14, M89, M213
et le noyau pour les dérivées
suivantes.
Noé possédait l’ADN-Y de base
qui a pu muter en ces autres
sous-groupes. Les divisions qui
sont issues de lui sont comme
suit :
Les Fils de Cham :
Le
Haplogroupe (Hg) A de l’ADN-Y
: M91 et les dérivées
subséquentes
A1 : M31,
A2 : M6, M14, M33, M49, M71,
M135, M141, P3, P4, P5, MEH1,
M196, M206
A3 : M32,
A3a : M28, M59
A3b M144, M190, M220
A3b1 : M51
A3b2 : M13, M63, M127,
A3b2a : M171
A3b2b : M118
Cet arbre est distinct des
autres Hgs B à R.
Ce groupe est trouvé
principalement au Soudan >45 %,
en Éthiopie >15 % et en Afrique
du Sud >30 %. De très petits
éléments de l’Hg A sont trouvés
au Cameroun, apparemment au
Maroc et parmi les Pygmées en
Afrique Centrale. La majorité de
ces autres sont ExE3b et B pour
les Pygmées et en Afrique du
Sud, ou E3b pour le Maroc.
L'Afrique est principalement
ExE3b et E3b avec le groupe B
réparti entre eux.
On voit aussi l'incidence de A
en Afrique avec le
Haplogroupe C
à sa racine, précédant un groupe
qui est resté en Afrique et une
branche qui a quitté l'Afrique
Le Haplogroupe B est composé de
M60, M181, puis :
B1 : M146
B2 : M182
B2a : M150
B2a1 : M109, M152, P32
B2a2 : M108a
B2a2a : M43
B2b1 : P6
B2B2 : M115, M169
B2B3 : M30, M129
B2b3a : M108b
B2b4 : P7
B2b4a : P8
B2b4b : MSY2a, M211
Le Haplogroupe B
apparaît en pourcentages plus
petits au Mali, au Cameroun,
parmi les Pygmées de l'Afrique
Centrale, en Afrique du Sud, au
Soudan et en Éthiopie. Les
Haplogroupes A et B
n'apparaissent pas de manière
importante à l'extérieur de
l'Afrique ou des
Afro-américains.
Le Haplogroupe C
Le
Haplogroupe C est un
Haplogroupe très intéressant. Il
est dérivé des groupes d'ADN
M168 et P9 et ensuite des
segments RPS4Y711 et M216 qui
forment le groupe C de base. Le
groupe C de base est trouvé
parmi les autochtones
australiens. Les divisions sont
:
C1 : M8, M105, M131
C2 : M38
C2a : P33
C3 : M217, P44
C3a : M93
C3b : P39
C3c : M48, M77, M86
C4a
C4b
Les groupes C se trouvent parmi
les gens suivants :
Aborigènes d'Australie >65% (C4)
Maoris >80% (C2)
Polynésiens français >55%
Samoans occidentaux >30%
Des pourcentages plus petits
sont trouvés en Irian Jaya,
Bornéo (Kalimantan), la Nouvelle
Grande-Bretagne et les
Philippines,
Des pourcentages plus petits
sont aussi trouvés en Chine et
au Japon.
Les pourcentages principaux sont
alors trouvés en Asie du Nord :
Mongols jusqu'à 60%
Bouriates jusqu'à 65%
Evenks jusqu'à 70%
Koriaks jusqu'à 35%
Altaïens jusqu'à 23%
Khazaks jusqu'à 42%
Ouzbeks jusqu'à 15%
Kirghizes environ 10%
Yakoutes 10%
Il y avait un élément qui s'est
déplacé dans les Amérindiens en
Alaska (disons environ 42 %) et
ensuite aux Chippewa ou Na-Dene
au Canada (+- 6 %) et aux
Cheyennes (>14 %) et aux Apaches
(>12 %) aux États-Unis. Il y a
un très petit pourcentage dans
les Esquimaux du Groenland
montrant un contact arctique
important.
C'est une question de grande
importance que nous avons été
capables de déterrer le point de
départ pour ces peuples et ces
tribus en Asie. La Caverne
Supérieure Choukoutien en Chine
(découverte en 1930 excavée par
Pei en 1938-1940) a été reconnue
comme étant la demeure d’un
groupe familial occupant les
mêmes strates et comprenant des
types de Mélanésiens,
d’Esquimaux et de Mongols.
Weidenreich a affirmé que les
restes de Wadjak étaient plutôt
les restes d’une femme adulte
dans la Caverne Supérieure
Choukoutien qu'avec n'importe
quels Méditerranéens jusqu'ici
(R. M et C. H Berndt,
Aboriginal Man in Australia
(Homme Autochtone en Australie),
Angus et Robertson, 1965, pp.
30, 32-33).
Précédemment, Huxley (1870)
avait classifié les autochtones
australiens comme étant
l'avant-garde des Dravidiens qui
ont quitté la Méditerranée et
l'Égypte et qui sont allés en
Inde et jusqu’en en Australie.
Il a considéré les tribus de
Dekkan Hill et de manière plus
précaire, les Égyptiens antiques
comme étant les seuls liens avec
les Australiens Autochtones
(ibid., p. 33). Il avait
classifié les hommes primitifs
en quatre types : Négroïdes,
Mongoloïdes et Xanthochroïdes
(de peau jaune) avec les
Australoïdes. Les Aborigènes
tasmaniens, il les a classifiés
comme une branche mélanésienne
des Négroïdes. Wallace (1893)
avait dit que les Australiens
n'étaient ni Négroïdes, ni
Mongoloïdes et les a classifiés
comme étant des Caucasiens
primitifs (ibid.). Cependant,
cela a été démontré comme étant
incorrect et les Autochtones
australiens sont descendants
directs de la division de
l'Haplogroupe C qui a quitté
l’Afrique
ou le Moyen-Orient. Ils se
composent également de deux
autres Haplogroupes, à savoir Hg
K des Mélanésiens et RxR1 de
base, qui est en fait un
Haplogroupe aryen de base à
partir duquel les Slaves R1a et
les Européens R1b sont
descendus.
Nous savons maintenant à partir
de l'ADN que les découvertes
dans la Caverne Choukoutien de
la même famille avec ces trois
types étaient en fait correctes.
L’ADN-Y des groupes tribaux
démontre que ces gens sont en
effet tous descendus des mêmes
lignes paternelles, et les
Australiens sont la structure C
de base qui est allée en
Australie de l'Afrique via le
Nord avec les Mélanésiens et les
Polynésiens développant le
Pacifique et les Mongols et ce
que sont devenues les tribus du
grand nord en allant dans l'Asie
du Nord.
Il y n’a donc aucun ADN publié
(connu de l'auteur) des
Australoïdes primitifs qui relie
les Autochtones Australiens
existants avec les premières
découvertes en Australie. Il y a
deux groupes principaux d’ADN-Y
en Australie et ils sont les
Haplogroupes C et K, avec un
premier Haplogroupe RxR1 >10 %
qui est trouvé en quantité
significative seulement au
Cameroun (>40 %) en Afrique et
avec des groupes plus petits en
Ouzbékistan et parmi les
Dravidiens et ils sont rares en
tous cas. Ce groupe RxR1
justifierait les liens à
l’Afrique et aux Dravidiens de
l’Inde. Cependant, c'est le seul
lien que les Aborigènes
australiens ont avec les
Dravidiens. Le groupe C de base
est un bien meilleur lien et
donc dans l’ensemble, les
Autochtones sont venus du groupe
qui a formé les groupes
africains qui a aussi formé les
groupes au Cameroun et dans le
Nord de l'Afrique orientale.
La grande majorité des Européens
sont R1a et R1b et pourraient
être considérés comme étant
descendus de ce lien d'ADN RxR1
primitif. Cependant, on
considère le
Haplogroupe RxR1
comme excluant toute origine
primitive pour les Aborigènes.
L'incidence de 25 % de l’Hg K
montre le mouvement des premiers
(proto-) Mélanésiens dans
l'Australie par le nord. Les
groupes C semblent être venus de
l'Afrique/Moyen-Orient dans
l'Asie Centrale. Cet aspect est
explicable bibliquement par la
division des Cuschites. Les
Cuschites occidentaux sont
entrés en Afrique par le Soudan
et les
Cuschites
orientaux sont allés d'abord en
Inde et ont ensuite continué en
groupes. Après l'invasion
aryenne de l'Inde en 1000 AEC,
ils semblent avoir quitté
l'Inde. Les Aborigènes se sont
séparés du groupe C de base sur
la côte sud asiatique,
probablement à cause de leur
existence nomade, et sont allés
en Australie probablement par
canoë. Les autres sont allés à
l’est et au nord en Asie et
ensuite dans le Pacifique.
Ainsi, on peut expliquer
l'origine africaine par une
division au Moyen-Orient plutôt
qu’un mouvement issu de l'Asie
et il y a donc une origine de
l’Asie centrale non seulement
pour cet élément des Aborigènes,
mais aussi pour les systèmes
entiers du groupe C des Mongoles
et du Pacifique. Les groupes R
de base sont reconnus comme
ayant été formés en
Inde/Pakistan plutôt qu'être
indicatifs d'une origine du
Moyen-Orient avec une migration
vers l'Afrique et l'Australie.
Nous regarderons cet aspect en
traitant de l’Hg R. Les divers
groupes linguistiques (8) pour
les Aborigènes indiquent environ
huit vagues en Australie pour au
moins trois systèmes tribaux
d’Hg et probablement une
migration plus diversifiée.
Il n'y a aucun Haplogroupe C en
Amérique du Sud et donc le
supposé lien entre les Maoris et
les Sud-Américains est un mythe.
L’ADN-Y prédominant en Amérique
est le
Haplogroupe Q avec le C y
apparaissant en nombre beaucoup
moindre et seulement de l'Alaska
vers l'Amérique du Nord centrale
dans les Na-Dene, Cheyennes et
Apaches
et dans les Mayas en Amérique
centrale.
On considère que le groupe C
pourrait être mal placé dans les
diagrammes et devrait peut-être
être relié de plus près aux
groupes K qui forment les proto-Mélanésoïdes
des Hgs K et M. Il pourrait
peut-être changer de place avec
le
Haplogroupe E, mais étant
donné les arguments de A et C
ci-dessus en Afrique, il
pourrait ne pas en être ainsi.
Le mouvement des groupes C,
c’étaient les groupes mineurs
avec Q qui sont allés dans les
Amériques. La sagesse
conventionnelle affirme qu'ils
se sont déplacés il y a
12-15,000 ans mais les
références de temps de la Bible
disent autrement.
Des Maoris ont affirmé à
l'auteur que les traditions des
canoës, qui relatent le
mouvement des tribus et lient
leur généalogie à ces canoës
arrivant en Nouvelle-Zélande,
permettent vraiment en fait une
possible origine africaine des
Maoris. Cependant, l'ADN
regroupant le C, avec
l'incidence de O, semble exiger
l'origine de l’Asie centrale et
le mouvement de la côte chinoise
coïncidant peut-être avec
l'expansion des Han.
Cependant, leurs liens
linguistiques malais indiquent
un mouvement au sud dans
l'archipel indo-malais où les
Maoris et les Aborigènes se
divisent en C4 et C2 par
l'isolement.
Il y a une trace d’ADN-Y (O)
asiatique de l'Est et du Sud-est
dans les Maoris, mais il y a des
éléments plus grands dans les
Samoans Occidentaux et, dans une
mesure légèrement moindre, parmi
les Polynésiens français. Cela
peut bien indiquer une influence
Malaise/Philippine sur les
tribus du Pacifique du système
C, mais une telle influence
n'est pas évidente en Irian Jaya,
en Papouasie-Nouvelle Guinée et
en Nouvelle-Bretagne où nous
pourrions nous attendre à les
trouver plus aisément. L'arrivée
des Maoris en Nouvelle-Zélande
est bien tardive et ils ont
remplacé deux civilisations
précédentes en Nouvelle-Zélande
dans les premiers siècles du
deuxième millénaire de l'ère
actuelle. La première était une
culture de type papoue, qui a
été forcée de migrer et la
deuxième semble être presque
celte dans ses structures en
pierre. Leur mouvement est ainsi
tout à fait distinct du
mouvement autochtone australien
précédent de C et de RxR1.
Le Haplogroupe D
Cet Haplogroupe est formé à
partir du groupe D et E à la
division YAP avec M145 et M203
comme la branche pour les deux
et M174 s’infiltrant dans le D :
D1 : M15
D2 : M55, M57, M64a, P37a, P41a,
12f2b
D2a : P12, P42
D2b : M116a
D2B1 : M125
D2b2 : M151
Cet Haplogroupe
est limité à deux peuples
d’importance. Ceux-ci sont le
Japon (>40 %) et le Tibet (>50
%). L'incidence de l’Hg indique
que les Japonais et les
Tibétains étaient jadis un
peuple occupant les Steppes
orientales et ils sont allés
vers le nord-est dans le Japon
et vers le sud dans les
montagnes du Tibet. La langue
des Japonais est
l’ouralo-altaïque et elle a
beaucoup en commun avec le
finlandais et le hongrois, le
turc, l’altaïque, le mongol, le
mandchou/Tungus, le vieux coréen
et les langues sibériennes du
nord (et anciennement avec le
basque). Donc, nous pouvons
supposer que le
Haplogroupe D
était jadis une tribu avec un
système de langue commun avec
ceux de l’Haplogroupe C et
d'autres identifiés ci-dessous.
L'autre Hg (O) pour les Japonais
et les Tibétains indique que les
Chinois s'étaient entrecroisés
avec eux au cours des siècles,
affectant sans doute le système
de langue et les coutumes des
régions de leur mouvement. Les
Japonais ont une incidence plus
haute de l’Hg C que les
Tibétains, peut-être dû à leur
exposition aux Mongols, aux
Bouriates et Koriaks. Cependant,
l'incidence de l'Autochtone
Aïnou peut aussi y être un
facteur en raison du
remplacement de la population
dans les invasions japonaises
postérieures.
L'autre incidence d’importance de D provient peut-être de l'influence du commerce des Japonais dans les temps anciens en allant à Sumatra et en Malaisie. L'incidence de plus grande importance est à Sumatra jusqu’à 10 %. L'incidence en Malaisie est moins de 5 %, comme le groupement R1a est là aussi et pour F, C et le M avec une incidence légèrement plus haute du K. Bornéo a moins de 5 % de groupe D aussi, aussi bien que les Ouïghours, Altaïques, les Mongols, les Kirghiz, les Ouzbeks et les Esquimaux Sibériens.
Le Haplogroupe E
Le Haplogroupe E se trouve en
Afrique pour l'essentiel. Il
continue la division Yap M145 et
M203 à SRY 4064 M96 et P29, qui
forme la base de E.
E1 : M33, M132
E1a : M44
E2 : M75
E2a : M41
E2b : M54, M85, M90
E3 : P2, DYS 391p
E3a : M2, P1
E3a1 : M58
E3a2 : M116b
E3a3 : M149
E3a4 : M154
E3a5 : M165
E3a6 : M10, M66, M156
E3b : M35, M215 (omis
de quelques diagrammes mais
contenu dans
Family Tree DNA 2005
Y-Chromosome Phylogenetic Tree
(Arbre généalogique ADN 2005
–L’Arbre Phylogénétique), voir
www.familytreeDNA.com)
E3b1 : M78
E3b1a : M148
E3b2 : M81
E3b2a : M107
E3b2b : M165
E3b3 : M123
E3b3a : M34
E3b3a1 : M136
Les groupes E sont trouvés en
Afrique avec une incidence plus
petite dans le Moyen-Orient et
l'Europe du Sud. Certains
spéculent l'incidence de E3a au
Royaume-Uni comme étant des
esclaves romains en
Grande-Bretagne, tandis que
d'autres les considèrent comme
des esclaves introduits en des
temps postérieurs. Les deux
opinions peuvent bien être
correctes.
ExE3b est le plus élevé au
Burkina Faso à bien plus de 90
%, peut-être 99 %.
ExE3b est élevé au Mali (60 %),
au Cameroun (45 %), parmi les
Pygmées (65 %) et en Amérique du
Sud (55 % approximativement). Ce
groupe a une faible incidence au
Soudan et en Éthiopie.
E3b est très élevé au Maroc
(approximativement 75 %) avec
une incidence de l’Hg J à
approximativement 10 %. Il y a
une incidence d’importance au
Mali (25 %), au Soudan (25 %) et
en Éthiopie (55 %,) en Afrique
du Sud (moins de 10 %). 15 % des
Arabes du Moyen-Orient et des
Persans sont E3b et environ 10 %
des Italiens et des Ibériens et
entre 5 % et 10 % des Allemands
et des Russes sont E3b. Jusqu’à
5 % des Géorgiens et des
Arméniens sont aussi E3b. Les
Juifs européens de l'Est sont à
25 % E3b. Ils peuvent bien avoir
été convertis au Judaïsme à
partir du Soudan et de
l'Éthiopie, vu qu’il y a des
éléments sémitiques dans les
deux régions.
Ils ont pu avoir fait partie de
la
multitude mélangée dans
l'Exode. Plus vraisemblablement,
beaucoup étaient les Cananéens
qui ont été épargnés et se sont
joints à Israël après
l’occupation sous Josué.
Nous examinerons ces aspects
quand nous regarderons les
Haplogroupes sémitiques
ci-dessous.
Ces Haplogroupes A à E sont les
fils de Cham. Nous tenterons
d'identifier leurs tribus à une
date ultérieure.
Tel que mentionné, le
Haplogroupe
C peut bien avoir été mal placé
et devrait probablement
apparaître après la position où
E se trouve dans l'arbre, au
côté de F du côté de ce qui est
appelé la
division
YAP. D et E sont tous deux
définis par une insertion sur le
Y que le D et le E partagent,
tandis que le C et le F n'ont
pas cette insertion. Ils
apparaissent plus fréquemment
ensemble dans les emplacements
tribaux.
Le Haplogroupe F
Le
Haplogroupe F est issu de la
division entre le Yap M145, la
division M203 pour le D et E et
la division RPS4Y 711 M216 pour
le groupe C. C'est un petit
groupe et il agit parfois comme
un fourre-tout parce que les
chercheurs n'ont pas fait assez
de tests pour déterminer le
groupe correctement. Il y a des
petits groupes F dans la
Géorgie/Arménie, la Perse,
l’Ouzbékistan, parmi les
Tartares Kazan et au Kazakhstan.
La conclusion est que la racine
de base de F est presque
disparue, mais les fils
prolifiques ont survécu et ont
prospéré, produisant ainsi les
groupes nationaux principaux et
les mutations qui ont découlé de
cette branche.
Cette branche détermine tous les
autres Haplogroupes de F à R.
Le F de base est P14, M89, M213.
En termes bibliques, tant Sem que Japhet ont passé ce Haplogroupe central à toute leur progéniture. Peut-être Cham l'a aussi passé à un de ses fils. Les diagrammes montreraient les fils de Cham comme étant très divergents.
Le puzzle de l’ADNmt
Avant
d'examiner davantage les
Haplogroupes (Hgs) de l’ADN-Y,
nous poserons un problème et
proposerons une solution.
Il y a 26
Haplogroupes ADNmt indiquant 26
lignées ADNmt féminines. Quelque
sept Hgs originaux ou "Ève"
féminines sont posés pour
l'Europe. Toutefois, lorsque
l'on examine l'arbre de l'ADNmt,
on trouve quelques dérivés
intéressants du groupe. Les
soi-disant “super-groupes” sont
en réalité seulement dans trois
groupes de base. En d'autres
termes, ils sont venus de trois
principales lignées par les
femmes. C'est ce que nous nous
attendrions à trouver si nous
supposons qu'il n'y avait que
trois femmes qui ont procréé
depuis l'Arche, à savoir les
épouses de Sem, Cham et Japhet.
Ces Haplogroupes sont tous des
descendants d'un seul
super-groupe féminin, à savoir
le Haplogroupe L. Donc, en
réalité, toutes les femmes sont
les descendantes d'une lignée
féminine, Hg L. Cela est super
L. Cette lignée s’est alors
divisée en L1, puis L2 et L3. La
ligne L3 a divergé et de L3 sont
venues les autres mutations de
l'ADNmt. Ainsi, toutes les
femmes provenaient d'une Ève
dont la lignée ADNmt était L.
Les groupes L
L1, L2 et L3 sont tous situés en
Afrique et sont les principaux
groupes presque exclusivement en
Afrique sub-saharienne.
Seulement de l'Éthiopie Nord
nous obtenons une grande
diversité de comptes rendus de
l’ADNmt. C'est la raison
fondamentale pour laquelle les
évolutionnistes affirment que
nous sommes tous venus
d'Afrique. La mise en place des
groupes d'ADN peut être vue dans
l’ouvrage de J.D. McDonald qui
les a regroupés par cartes d’Haplogroupes
Y et ADNmt disponibles au
http://www.scs.uiuc.edu/~mcdonald/WorldHaplogroupsMaps.pdf.
Les
super-groupes M et N devaient
ensuite diverger ou muter. D'un
point de vue biblique, nous
pouvons débattre facilement que
L a été formé avec Ève et les
autres groupes étaient des
divisions pré-Déluge qui sont
venus sur l'Arche. Ainsi, nous
pourrions soutenir à juste titre
que L, M et N sont venus sur
l'Arche dans le récit biblique
accepté. Il est également
possible que la subdivision
super-groupe R puisse être venue
sur l'Arche, en fonction du
nombre de femmes. Tous les
Haplogroupes ADNmt sont des
subdivisions de L, alors M et N,
puis R, qui lui-même est une
mutation de l’Hg N. Ainsi, le
seul argument entre le récit
biblique et l'ADN de la science
moderne est la supposition que
les modèles mathématiques
exigent une période beaucoup
plus longue pour muter que la
chronologie de la Bible. Cette
hypothèse est basée sur la
prémisse que l'ADNmt ne force
pas la mutation du Génome
Humain. Cette supposition est
maintenant révélée comme étant
fausse. La déclaration de la
Bible que “la mort est venue par
le péché” est apparemment
manifeste comme un état du
système de l'ADN humain, comme
nous l'avons vu précédemment.
Comme le montre l'étude de
l'Institut Pasteur, l'ADNmt
cause une mutation où les
dommages ont eu lieu par la
maladie, et aussi la radiation,
comme nous sommes en train de le
découvrir. Nous mutons
progressivement dans notre ADN à
travers l'influence directe de
l'ADN Mitochondrial sur le
Génome Humain. Nous avons
commencé parfaits avec Adam et
nous nous affaiblissons au fil
du temps. Nous n'avons pas
évolué, nous avons dégénéré.
Notre ADN s’est muté à
l'exposition de l'ADNmt dans ses
variantes formes. Il s'ensuit
comme une question de logique
que l’ADNmt divers doit causer
de nouvelles mutations dans le
Génome Humain qui affectent le
Chromosome Y.
Ainsi, à
partir de l’Hg L d'origine nous
obtenons Hg M et Hg N. Ces deux
groupes sont indépendants des
mutations directes de l’Hg L.
Donc on peut
supposer qu’Ève a produit la
ligne L et les trois femmes de
Sem, Cham et Japhet sont au
moins les trois groupes L, M et
N. Il peut y avoir eu de
nouvelles divisions, étant donné
le fait que Noé a peut-être eu
des filles non mentionnées, et
leur lignée de l'ADNmt peut
avoir été L, ou M ou N. Elle a
peut-être même été R, si nous
supposons que la lignée L
entière est entrée par la femme
de Cham, puisque la lignée L est
presque confinée aux tribus
sub-sahariennes. Nous devons
également tenir compte du fait
qu’Ève avait la peau sombre et
le fait qu’Adam signifie
celui qui était rouge. Ainsi
la capacité de développement de
la couleur de la peau est une
caractéristique originale de la
création humaine. L'explication
de la diminution de mélanine, ce
qui provoque la pigmentation,
c'est que plus vous vous
éloignez de l'équateur plus
votre peau doit être claire pour
survivre. Vous obtenez moins de
vitamine D avec une augmentation
de la mélanine. Ainsi, vous avez
besoin de moins de capacité à
absorber la vitamine D, où le
soleil est le plus grand, et la
protection de plus de mélanine,
mais une plus grande capacité à
absorber la vitamine D et moins
de protection où le soleil est
diminué. Ainsi, les descendants
de Noé sont simplement devenus
plus clairs en mélanine
lorsqu’ils se sont déplacés plus
loin au nord.
Les épouses de
Sem et Japhet étaient des
subdivisions de L, peut-être M
et N ou peut-être aussi R.
Les deux
groupes M et N ont formé des
sous-groupes suivants :
M a produit
trois subdivisions :
Sous-groupe M,
y compris
C et Z, qui se
sont séparés l’un de l'autre, et
D et G ;
Subdivision E
; et
Subdivision Q
On pourrait
donc aussi en déduire que les
femmes des fils de Noé ont été
prises à partir de l’unique
lignée familiale préservant la
pureté dans les générations dans
la ligne féminine aussi. La
scission L2 et L3 peut provenir
de la structure familiale
d'avant le Déluge. Les filles de
Noé et les femmes des fils
auraient pu porter les trois
subdivisions L et les
sous-groupes de base de M, N et
peut-être R.
Il est par
conséquent possible que les
femmes de l'Arche – même s’il
n'y avait que les filles de Noé
et les épouses de Sem, Cham et
Japhet – auraient pu facilement
contenir la base de la diversité
moderne de l’ADNmt. Nous ne
serions pas surpris de trouver
une telle diversité dans une
famille de fils mariés, même
aujourd'hui. En Palestine, en
Égypte et au sud du Pakistan, il
est courant aujourd'hui de
trouver ces groupes.
Le super-groupe M
M est trouvé en quantités et en pourcentages plus grands
parmi les Indiens et les
Pakistanais. De là, le groupe
s’étend vers le nord et vers
l'est aux Mongols et aux Chinois
Han et jusqu’au Japon et Taïwan.
Les divisions de M en C et D
sont arrivées dans les steppes
orientales parmi les Hans et les
Mongols et les Japonais. Les
divisions C se sont produites
parmi les Mongols et les
sous-groupes. Les Evenks sont
principalement C avec seulement
de petits pourcentages de M, D
et U. Les Bouriates de la
Sibérie Orientale Centrale sont
C et D avec un peu de L3 et
ensuite la mutation G. Cette
mutation est transportée vers
l'est dans les Nivikhs et est la
plus prolifique parmi les
Itelmen de la Sibérie Orientale
et s’étend dans les Chukchi de
la Sibérie d'Extrême-Orient.
La
mutation de M en Hg E s’est
produite parmi les Taïwanais
Autochtones et au Kalimantan,
étant prolifique au Sabah. La
variante Q est arrivée en
Papouasie-Nouvelle-Guinée. Donc
la susdite variation d’ADNmt est
arrivée à mesure que les tribus
se sont déplacées
progressivement vers l'est à
partir des Indous Cusch vers la
Mongolie, la Chine, l’Asie du
Sud-Est et la Mélanésie.
Ceci a été la progression
originale de l’ADNmt du
super-groupe M après le Déluge.
Le super-groupe N
Le super-groupe N est maintenant
trouvé surtout en Asie Orientale
et Centrale et parmi les
Autochtones australiens. Le
groupe N s’est divisé en :
Le sous-groupe N, qui a inclus
les Haplogroupes I et W ;
La subdivision A et X ;
La subdivision Y ; et
La subdivision R.
Des concentrations de la base N
sont parmi les aborigènes
australiens approximativement
dans la même distribution
puisque l’Hg C de l'ADN-Y est
trouvé là. Ce fait indique que
ces gens sont venus en Australie
probablement de la distribution
orientale des Cuschites et sont
restés isolés. Leur ADN de base
n'a pas subi de mutation par
d'autre intrusion de
mitochondrie. Leur structure de
base ne donne pas d’arguments
pour une longue occupation, mais
plutôt un système d'ADN isolé
qui n'a souffert d’aucune
mutation forcée par les mariages
mixtes avec d'autres
Haplogroupes de l’ADNmt.
La structure P en Australie peut
être entrée avec l’Hg K et RxR1
de l'ADN-Y des Mélanésiens. Il y
a de fortes concentrations des
deux Hg P et Q de l’ADNmt avec
un peu de M. Il y a un très
petit pourcentage d’ADNmt M en
Australie avec un petit
pourcentage similaire de ADNmt N
en Papouasie-Nouvelle-Guinée.
Cela indiquerait un échange
occasionnel, probablement de
femmes capturées. Il y a un
argument que l'ADN-Y K et l’ADNmt
P peuvent avoir été là avec les
Papous quand les Autochtones
tasmaniens sont venus et ont en
fait précédé l’ADN-Y C et l’ADNmt
N autochtones australiens. Cela
est incertain mais les
Tasmaniens étaient d'un type
papou comme étaient les Morios
qui ont précédé les Maoris en
Nouvelle Zélande.
La structure de l’ADNmt N est aussi parmi les Japonais,
les Chinois Han, les Taïwanais
Han, les Altaï, les Ouzbeks, les
Komi, les Persans, les Turcs et
aussi en Russie du Sud. Le
super-groupe N s’est divisé en
sous-groupes N, I et W.
Le sous-groupe I est quelque peu
rare et apparaît comme un
pourcentage significatif, bien
que petit, parmi les Kurdes et
en Turquie probablement à cause
des Kurdes là. C'est un petit
pourcentage en Égypte et
continue ensuite en Ibérie, en
France, au Royaume-Uni, en
Scandinavie et en Islande. Il
semble que la division de N et
de I soit arrivée au
Moyen-Orient avant que les
groupes N ne se soient déplacés
à l'est et que l’Hg I se soit
déplacé à l'ouest. W se trouve
aussi parmi les Kurdes comme le
sont U et K et HV et la
subdivision H. Donc, ces deux
divisions et les sous-groupes
sont représentés là, mais le
parent Hg R n'est pas
représenté. Les groupes N, I, et
W sont tous présents en
Palestine/Égypte. L’Hg I de l’ADNmt
est probablement venu du
Moyen-Orient avec les
Anglo-Saxons ou les premiers
mouvements celtiques, peut-être
même après la chute de Troie au
11ème siècle AEC.
Les Haplogroupes ADNmt A et X
sont prédominants parmi les
Indiens d’Amériques. Étant une
subdivision, Hg X est arrivé
quand ils se sont déplacés dans
le Nord-Est de l'Amérique du
Nord.
Y est une mutation qui est
arrivée au Nord de l'Asie et
elle est trouvée parmi les
Nivikhs.
Le super-groupe R
Le Haplogroupe R est trouvé
parmi les Thaï dans les plus
grands pourcentages, en Inde, au
Sud du Pakistan et en
pourcentages plus petits parmi
les Chinois Han et les Hazara.
Le super-groupe R s’est divisé
comme suit :
B ;
F ;
HV, qui s’est divisé en H et V ;
P ;
Les subdivisions J et T ; et
U, d’où est issu K.
Alors que l’Hg R est trouvé
parmi les Thaï et d'autres comme
ci-dessus, la sous-division B
est plus révélatrice. B est
trouvé parmi les Chinois Han et
en pourcentages plus grands en
Chine du Sud avec de plus grands
pourcentages parmi les Taïwanais
Autochtones, mais présents aussi
parmi les Taïwanais Han et les
Japonais. Le Haplogroupe B s’est
déplacé vers le sud en
Papouasie-Nouvelle-Guinée et se
trouve dans les concentrations
les plus élevées parmi les
Polynésiens, étant plus de 90 %
de leur ADNmt. Le sous-groupe B
est alors trouvé à travers le
Pacifique dans les
concentrations les plus grandes
sur la côte ouest des Amériques
centrale, du Sud et du Nord
indiquant que le mouvement était
de l'Asie Centrale à l'Asie du
Sud-Est vers la Polynésie
jusqu’aux Amériques, mais
l'ADN-Y dans les Amériques n'est
pas C mais Q. C'est seulement C
avec Q en Alaska et ensuite
jusque dans l'Amérique du Nord
Centrale. Cet itinéraire de C
doit aussi avoir été à travers
le nord ou, les tribus de
l’ADN-Y Q ont pris les femmes
des hommes de l'ADN-Y C et les
ont anéanties. Cela peut
correspondre aux récentes
découvertes d'un mâle de base
d’ADN-Y C – identifié comme un
Autochtone australien sur la
Côte ouest de l'Amérique du Nord
et antérieur à n'importe quelle
autre découverte jusqu'à présent
– qui semblent indiquer cette
migration.
La présence de groupes A, X et D
rassemblés indique une traversée
du Pacifique de l'Asie du Nord
avec la division A de l’Hg N qui
arrive à Taïwan et au Japon et
se propage au nord jusqu’au
Chukchi de la Sibérie
d'Extrême-Orient et à travers
les Alleutiers en deux groupes.
Le groupe du nord devait devenir
les Esquimaux de l’Alaska et le
groupe du sud devait devenir les
Amérindiens de l’Alaska. Les
groupes du nord semblent avoir
été les principaux migrateurs du
nord formant l’ADNmt des Na-Dene
(Chippewa) (principalement de
l’Hg A). Le groupe A est allé à
l'Est et au Nord et jusqu`au
Groenland avec les groupes D
continuant en Amérique du Sud
avec A, C et B aussi bien que
dans la région que sont les
États-Unis du Sud-est. Les
hommes Q avaient donc quatre
lignées de femmes avec eux
incluant la variation
postérieure de l’ADNmt U étant
trouvé en Amérique du Nord, mais
aussi dans la région qu'est
aujourd’hui le Chili. Ainsi
l’établissement des Amériques a
été en retard avec les mutations
de l'ADN-Y et de l’ADNmt étant
des secondes aux dernières
formes.
Le groupe HV a subi une mutation
de R au Moyen-Orient et tous
deux R et HV sont trouvés en
Perse et au Pakistan du Sud. La
variante Hg H est trouvée parmi
les Ouzbeks et les Mansi, les
Hazara et les Komi dans les
Steppes Centrales à travers
l'Europe jusqu’en Islande. H
forme la structure d’ADNmt
principale des Européens de
l'Ouest. Il est aussi trouvé en
Palestine, en Égypte et en
Afrique du Nord-Ouest jusqu’au
Maroc. La variante Hg V est dans
des quantités plus petites sauf
parmi les Saami de la
Scandinavie du nord où les
pourcentages sont inversés de
façon dramatique. Les
pourcentages de J, T, U et K de
la sous-division R sont trouvés
répartis de façon semblable à
travers l'Europe à l'exception
des Saami et des Kurdes où U est
prédominant sauf la variante K
étant trouvée parmi les Kurdes.
La variante K est absente dans
les Saami indiquant qu'ils n'ont
pas vu cette variation, se
séparant des autres groupes U
avant qu’elle ne se produise.
Les autres Tribus du Nord en
Russie, des Kets et des Komi ont
un nombre significatif de U mais
elles ont aussi des variations F
ou T, que les Saami n’ont pas.
Les Saami ont aussi un certain
pourcentage de D aussi bien que
H et un peu de Z.
Les divisions de l’Haplogroupe
ADNmt progressives sont
présentées dans le tableau
ci-attaché.
Arbre Simplifié des Haplogroupes
Mitochondriaux
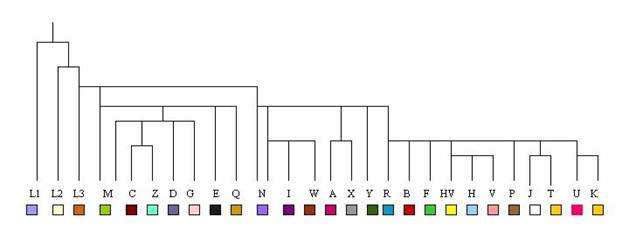
Notez que, contrairement aux
Haplogroupes du Chromosome Y,
ceux pour l'ADN Mitochondrial ne
sont pas liés de façon simple à
l'ordre alphabétique. Cela est
dû à la nature historique de
leur définition. Les groupes
indigènes d'Amérique A, B, C et
D ont été mis en place par ordre
alphabétique. Ils ne sont pas,
comme on le voit ci-dessus,
étroitement liés à l'arbre, M, N
et R sont appelés "superhaplogroupes"
comme indiqué. Notez que M et N
sont en fait un peu hétérogènes.
Conclusions
Il
n'y a donc rien dans les variations d’ADNmt et
les Haplogroupes pour exclure l'histoire de la
Bible et le récit de la Genèse qui sont
accommodés par ou accommodent les avances
scientifiques que nous trouvons ici.
Les
Haplogroupes ADNmt L, M, N et peut-être R
étaient présents dans les femmes sur l'Arche.
Les mutations se sont produites au fur et à
mesure que chaque groupe s’est déplacé en dehors
du Moyen-Orient et s’est reproduit avec des
tribus et des familles en déplacement au fil du
temps.
Ensuite, nous retournerons à la structure d'ADN-Y pour essayer d'identifier les tribus et les nations impliquées et de les placer dans le récit de la Genèse.
Les groupes de l’ADN-Y
Tous les autres groupes de l’ADN-Y à partir de G
à R2 sont dérivés d'un Haplogroupe central F. Ce
groupe F est au cœur de lignées connues à la
fois de Sem et de Japhet. Nous allons commencer
par la Lignée F, qui est P14, M89, M213. Cette
ligne est la lignée de base pour G, H, I, J et
K.
Nous savons que les lignées de peuples
sémitiques connus sont G, I et J. Il y a aussi
quelques lignées des lignées E3B africaines ou
chamitiques, avec quelques-unes R1a et R1b. Ces
lignées avec G s'étendent aussi à la Turquie, la
Géorgie/Arménie et l'Italie.
La sagesse conventionnelle indique les Arabes du
Moyen-Orient comme le Haplogroupe J, et le
sacerdoce juif d’Aaron, qui a une lignée claire
identifiée à Sem, est à J2. Cette division
d’Haplogroupe identifie également le clan Buba
de la tribu des Lemba du Zimbabwe en tant que
sacrificateurs d'Aaron et ils ont été séparés du
reste de Juda/Lévi pendant 2,500 ans. Ainsi, la
division J2 est au moins aussi ancienne que
cette séparation.
Il y a aussi un nombre important de divisions
dans le Judaïsme qui montrent que le Judaïsme
est une religion et non une lignée d’un seul
Haplogroupe.
Par exemple, tandis que le sacerdoce d'Aaron est
identifié comme J2, la structure lévitique de
Juifs ashkénazes est 52% R1a1, qui est une
lignée japhétique identifiée survenant en Russie
et dans les Steppes centrale et orientale et
parmi les Aryens de l'Inde. Elle est slave. En
outre, 25% de tous les Juifs d'Europe orientale
sont E3b, ce qui indique une origine hamitique.
Une hypothèse concernant l'origine E3b est que
les Égyptiens se sont croisés avec les
Israélites. Une autre est que la Multitude
Mélangée impliquait des E3bs, car il y avait
quelque deux millions d'Israélites et environ
600.000 de la Multitude Mélangée. Cela
constituerait 25%. Cependant, quand nous lisons
les comptes rendus génétiques des Milésiens ils
prétendent avoir fait partie de la multitude
mélangée aussi dans l'Exode. Ils sont R1b.
Ainsi, l'explication doit impliquer des
conversions au Judaïsme postérieurs à partir de
lignées hamites. Nous trouvons celles-ci dans
l'occupation de Canaan sous Josué.
Canaan était un fils de Cham et un certain
nombre de sous-tribus ont été épargnées et ont
rejoint Israël dans certains cas par subterfuge.
Les conversions éthiopiennes ont également de
l'importance et nous avons donc une ample
explication pour les lignées E3b à 25% des
Ashkénazes. Les Amoréens et les Hittites du Sud
sont également entrés et se sont reproduits avec
Israël. Il existe également des niveaux
importants de E3b en Syrie, en Turquie et chez
les Grecs jusqu'à 30%. Les Grecs sont jusqu'à
30% de l’Haplogroupe J ou de sémites connus
également avec jusqu'à 20% de l’Hg I aussi, ce
qui est aussi un Haplogroupe répandu à la fois
chez les Sémites et les Européens connus. Nous
allons traiter de ces aspects plus tard.
L’ADNmt Ashkénaze
Un autre aspect important est celui de l'ADNmt
des Juifs ashkénazes. L'ADNmt des Juifs
ashkénazes est considéré comme venant du
Proche-Orient, mais nous devons examiner la
structure. Comme nous l'avons vu avec l'ADNmt,
tous les autres groupes sont venus de l’Hg L3,
qui s'est scindé en deux groupes principaux M et
N. De N est venu R et les divisions subséquentes
de N et R. N s’est divisé au Moyen-Orient et a
formé le sous-groupe Nu de N (N1), I et W. Nous
savons que I s’est produit parmi les ancêtres
des Kurdes dans ce qui est maintenant le
Kurdistan et le pays montagneux sur les zones
frontalières de l'Irak et de l'Iran ou la Perse,
où les Mèdes (qui sont les fils de Madaï) ont
vécu. Les autres groupes de ce sous-groupe ont
formé N1, etc.
10% de l’ADNmt des Juifs ashkénazes sont N1b. Ce
type ne diffère que par une seule transition du
N1b ancestral (145-176G-223). C'est peut-être
parmi les plus anciens de l'ADNmt à l'exception
de L2. Cet ADN L2 comme nous l'avons vu, est une
émanation de l'ADNmt L primitif. Il est
d'origine hamitique et surtout maintenant
subsaharien et éthiopien. Il est peut être venu
des Cananéens ou de la multitude mélangée. La
première femme de Moïse était une Éthiopienne
aussi et les Cananéens peuvent bien avoir eu cet
ADN. Il y a aussi un type proche-oriental de M1,
qui est la subdivision primitive du groupe M.
Les autres sont toutes les subdivisions de R, et
qui constituent 60% de l'ADNmt ashkénaze.
Les types d'ADNmt du Proche-Orient chez les
Juifs ashkénazes sont N1b, H, J, K, L2, preHV,
U7, M1, U1B (cf. Behar et al.,
Differential Bottleneck Effects in the mtDNA
Gene Pool of Ashkenazi Jewish Populations,
pp. 8,9,19 ; voir aussi
MtDNA Evidence for a genetic
bottleneck in the early history of the Ashkenazi
Jewish population, European Journal of Human
Genetics,
2004, pp. 1-10).
Il y a un faible pourcentage de l'ADN
mitochondrial comme U5a et V, qui sont
considérés comme venir de mélange européen
(Behar et al., P. 10). Toutefois, V est
prédominant chez les Saami avec des pourcentages
mineurs à nettement moins de 10% parmi les
Européens.
La dernière division est celle de U, qui s’est
scindée pour former K.
32% de l’ADNmt de tous les Juifs ashkénazes sont
K, composés de 4 sous-types principaux. Parmi
les non-juifs le pourcentage le plus élevé est
parmi les Druzes à 16%. L’ADNmt des Juifs
ashkénazes le plus élevé qui suit est H à 21% et
duquel 7% est H ancestral. L’ADNmt des Juifs
ashkénazes britanniques est le haplotype [ou
haplogroupe] modal
CRS. Ainsi, on peut supposer que la division
initiale des HV s’est probablement produite au
début de la population de ces femmes (début
d'Israël ?). J1 est le prochain à 7%. (Behar op.
cit.)
Ce qui est important à cet égard c’est que H
constitue 21% de l’ADN mitochondrial des Juifs
ashkénazes et cet ADN est la séquence d'ADNmt
principale de la population européenne dans son
ensemble à un peu moins de 50% de l'Islande,
jusqu’à 60% des Îles britanniques, et 65 %
d'Iberia et de 50% de la France. Il diminue
lentement à mesure que les populations vont à
l'est. Il tombe à 25% chez les Turcs et la
Palestine/Égyptiens et diminue davantage chez
les Kurdes, les Perses, les Pakistanais et les
Hazara du Sud, avec une augmentation de plus de
22% parmi les Ouzbeks et jusqu'à 30% dans le
nord de la Russie parmi les Komis.
Il est donc raisonnable de supposer que les
mêmes femmes qui étaient la base des Européens
de l'Ouest ont été la base pour beaucoup de
femmes ashkénazes, et que les mutations se sont
produites au sein de ces tribus.
La mutation fondamentale de U à K semble avoir
eu lieu au Moyen-Orient dans la région du nord
d'Israël et le Liban, et comprenait une grande
partie des femmes qui ont formé l'ascendance des
Druzes. Ils sont les ancêtres de 32% des Juifs
ashkénazes. Les autres personnes avec une forte
proportion de K sont les Kurdes desquels U forme
la majeure partie. U forme une partie importante
de tout l'ADNmt européen. Il est plus élevé chez
les Saami. HV, H, U, V et K sont les principaux
groupes de tous les Européens avec I et J. H
constitue plus de 50% de tous les Anglo-Celtes,
les Scandinaves et les Français et est encore
plus important en Iberia. La différence
significative entre les Ashkénazes et les
Européens, c'est que la division K est plus
élevée chez eux qu’elle ne l’est en Europe et
ainsi le mouvement de K peut avoir eu lieu plus
tard en Europe ou a été moindre dans sa mutation
à partir de U là. Il est probable que ces
divisions soient survenues à partir de la
rupture de l'Alliance des Parthes et le
mouvement de la Horde en Europe sous Odin, à
compter de la fin du IIème siècle EC.
Les deux groupes ont probablement formé la même
Horde scythe composée de Celtes et d’Israélites
et de Juifs de l'Alliance hittite. La Horde
parthe et scythe s’est déplacée vers l'Europe et
les autres tribus ont formé la Horde Khazar et
est devenue l'Empire khazar, se convertissant au
Judaïsme ca. 740 EC, jusqu'au XIème
siècle et leur défaite par les Russ et le
mouvement postérieur en Europe avec les Huns.
Cela explique pourquoi l'ADN lévite
non-sémitique dans les Lévites ashkénazes (qui
est la majorité de son ADN-Y) est R1a1, qui est
slave plutôt que celte.
Il faut aussi rappeler que l'identité juive
depuis l'époque talmudique ne suit pas
rigoureusement le système de la Bible et a été
acquise par filiation maternelle d'une femme
juive ou par conversion rabbinique autorisée
(voir aussi Behar et al,.
Multiple Origins of Ashkenazi
Levites: Y Chromosome Evidence for Both Near
Eastern and European Ancestries,
Am. J. Hum.
Genet. 73:000-000, 2003). Il est considéré que
l'ADN est venu de la conversion des Khazars, et
peut-être autant que 50 hommes initialement
inscrits comme Lévites de la Horde Khazar slave.
Il semble y avoir également un événement d’un
mâle unique qui s’est également considérablement
élargi après le mouvement en Europe (ibid.).
Cohenim et les Lévites comprennent chacun
environ 4% de la population juive. Selon l'Encyclopaedia
Judaica (1972), les Gentils peuvent devenir
des Israélites, mais pas des Lévites ou Cohenim
(cf. ibid.). Cette règle évidemment n'a pas été
suivie dans la conversion des Khazars. Dans les
haplotypes [ou haplogroupes] R1a1 des Lévites ashkénazes les
haplotypes microsatellites sont fortement
regroupés autour d'un haplotype modal
(16-12-25-10-11-13), qui comprend 74% des
Lévites ashkénazes dans le Haplogroupe et 38% des
Lévites ashkénazes en général. Le haplotype est
réparti uniformément en général (Behar et al.,
ibid., voir également le Tableau 4).
Les Juifs tentent de restreindre les origines
des Ashkénazes à la petite zone du nord-ouest de
l'Europe du Nord-est de la France à l'Allemagne
du Nord. Ils ont fait valoir une origine dès le
6ème siècle. Cette région à cette
époque était l'emplacement central de la Horde
parthe comme elle a rompu et s’est déplacée en
Grande-Bretagne, en Scandinavie, en France et en
Espagne et en Italie. Il est probable que
certains éléments juifs de la Horde s'y sont
installés ou ont suivi la Horde plus tard. Le
problème est que les définitions précédentes,
qui ont vu le yiddish se développer à partir du
10ème siècle, sont que la langue
Ashkenazi était en apparence un langage basé sur
les lettres hébraïques mais avec des mots
empruntés au lexique allemand. Cependant, elle
est en effet une langue slave relexifiée
(Wexler, 1993, cf. ibid.). Elle est liée au
sorabe et au bélarus. Cette hypothèse est
corroborée aujourd'hui par la majorité de
l’ADN-Y slave des Lévites ashkénazes. Elle est
maintenant considérée comme sorabe qui a reçu
des lettres hébraïques et une lexicographie
germanique. L’ADN-Y R1a1 concerne également
l’ADN-Y sorabe et bélarus. Ces tribus pourraient
alors être considérés comme les fils d'Ashkenaz
et une partie de la Horde Khazar, qui est donc
manifestement slave.
Le terme séfarade, lorsqu'il est appliqué
aux Juifs, était lié aux Juifs d'Espagne avant
l'expulsion de 1492, et où les Juifs étaient
aussi en Grande-Bretagne. Cependant
l'utilisation actuelle l'applique à tous les
Juifs d'Afrique du Nord et du Proche-Orient qui
suivent le rite séfarade d’adoration.
L’ADN-Y des Cohenim ou sacrificateurs lévites,
par opposition aux Lévites et Israélites, par
ailleurs, est un haplogroupe spécifique. Les NRY
(ADN-Y) des Cohenim Ashkénazes et Sépharades
sont manifestement plus liés les uns aux autres
qu’aux autres Israélites et non-Israélites
(Skorecki et al., 1997 ; Thomas et al., 1998,
cf. ibid.). Le motif de leur ADN provient
principalement de différences dans la fréquence
de l'Haplotype Modal Cohen (CMH), défini par six
microsatellites à mutation rapide (Thomas et
al., 1998), et un groupement d'haplotypes
étroitement liés au sein d'un seul Haplogroupe
(J2) défini par des polymorphismes à événement
unique avec une mutation lente. Les chromosomes
appartenant à ce haplotype et son groupement lié
ont été trouvés à fréquence élevée chez les
Cohenim, mais à une fréquence beaucoup plus
faible chez les
“Israélites”.
Le motif de groupement a été jugé conforme à la
descendance d'un ancêtre commun qui a vécu il y
a entre 2100 et 3900 ans. Maintenant, ce sont
les conclusions fondées sur des modèles
mathématiques qui ne reconnaissent pas la
mutation rapide de l’ADN-Y due à l'exposition de
l’ADNmt. En effet, ils seraient limités dans ce
haplotype aussi à cause des limitations sur le
mariage des Cohenim à l'extérieur de Lévi.
Le maximum plus tôt possible pour la formation
du CMH serait 1900 AEC et ainsi Abraham ne
pouvait pas avoir possédé ce haplotype. De plus,
la date inférieure, qui est la plus probable,
indique que le CMH a été fixé aussi tard que la
fin de la période du Temple en 100 avant notre
ère. Il pourrait bien être le résultat de
l'interaction en Judée des Édomites qui ont été
convertis au Judaïsme sous Jean Hyrcan ca. 160
AEC et les Juifs eux-mêmes là-bas. Ce fait peut
expliquer pourquoi le CMH n'est pas très répandu
en dehors des populations rabbiniques quand on
sait pour un fait absolu qu'il y avait une
population Lévite étendue entre les divisions du
sacerdoce en Israël. Toutes sauf trois de ces
vingt-quatre divisions sont reparties avec la
Horde Israélite quand elle a été emmenée en
captivité par les Assyriens au huitième siècle
avant notre ère et amenée au-delà de la rivière
Araxe dans ce qui est devenu la Parthie et la
Scythie. Il devrait y avoir un nombre important
de l'original ADN-Y Lévite/Cohenim avec la Horde
où qu'elle soit. L'apparition du CMH dans le
clan Buba des Lemba au Zimbabwe indique soit que
la mutation dans le CMH et J2 s’est produite à
l'époque de la Captivité babylonienne lorsque
les Lemba revendiquent qu’ils ont été séparés et
après la division d'Israël dans les dix tribus
et les deux tribus de Juda et une partie de Lévi
et quelques-uns de Benjamin, ou leur séparation
s’est faite plus tard et près de l’an 160 AEC
lorsque certains sacerdoce se sont déplacés en
Égypte et ont construit le Temple après
l'apostasie d'Antiochus Épiphane ou à la
dispersion de la Judée dans la période ca. 135
de notre ère après l'édit d'Hadrien et/ou
pendant la Révolte de Bar Kochba. Le
développement du F sémite des Patriarches à ses
variantes (par exemple G, I et J) peut très bien
avoir pris cinq cents ans ou plus au
Moyen-Orient et en Scythie et avoir eu lieu au
cours du premier millénaire avant notre ère
ou dans le premier
siècle de notre ère. La dernière date est la
plus probable pour la dispersion, c'est-à-dire,
sous Hadrien.
Nous pouvons donc en déduire que le Haplogroupe
J n'était pas le haplotype d'origine sémitique.
Les Sémites viennent donc de plus d'un
Haplogroupe standard dans la descendance
d'Abraham. Le Haplogroupe d'origine sémitique
était F divisé I en et J à tout le moins, et
sans doute G, I et J du Moyen-Orient, en
Parthie/Scythie et en Europe. Un examen de cette
hypothèse peut être conduit par référence aux
résultats de Behar et al.,
Multiple
Origins of Ashkenazi Levites
ibid., Tableau A. Dans ce tableau, les Lévites
ashkénazes ont été testés à partir de 60 lignées
mâles de plus de neuf pays. Ces pays sont
l'Autriche-Hongrie (AH) x 10, la Biélorussie
(Bel) x 4, la France (Fra) x 6, l’Allemagne
(Ger) x 10, la Lituanie (Lith) x 8, le Pays-Bas
(Neth) x 5, la Pologne (Pol) x 7, la Roumanie
(Rom) x 4, et la Russie (Rus) x 6.
Des 60 lignées d’ADN-Y testées seulement huit
étaient des Sémites réels. Les autres résultats
ont été les suivants :
Lignées hamitiques connues 12
DE (xE3) x 1 (Fra)
E3b x 11 (4 AH, 1 x Bel, 1 x Lith, 2 x Rom, 2 x
Rus, 1 x Ger).
Lignes japhétiques présumées 40
K (x L, N, O, P) x 1 (1 x Ger).
N (xN3) x 1 (1 x Lith).
Q x 1 (1 x Rus).
R1 x 1 (1 x Ger).
C'est une lignée très rare trouvée chez deux
Arméniens précédemment.
R1a1 x 31 (4 x AH, 3 x Bel, 3 x Lith, 6 x Pol, 1
x Rom, 1 x Rus, 4 x Fra, 7 x Ger,
22 x Neth).
R1b x 5 (2 x AH, 1 x Rus, 1 x Fra, 1 x Neth).
Les lignées sémitiques sont les suivantes et
contribuent à démontrer le point de la
divergence des lignées sémitiques discutées
ci-dessus.
F (x G, H, I, J, K) x 1 (1 x Rom)
I x 1 (1 x Neth).
J (x J2) x 2 (1 x Lith, 1 x Rus).
J2 x 4 (2 x Lith, 1 x Pol, 1 x Neth).
Les implications de ces échantillons d’ADN-Y
extrapolent passablement. Il y a seulement 4
lignées qui partagent les mêmes lignées J2 comme
le CMH. Seulement 6 partagent le Haplogroupe J.
Ainsi, seulement 10% des Lévites ashkénazes
testés sont reconnus comme de l’ADN-Y
juif/israélite direct connu. F et I représentent
25% de l’ADN-Y sémitique Lévite testé.
Sous la compréhension actuelle, les chiffres
impressionnants sont que deux tiers de l’ADN-Y
des Lévites ashkénazes ne sont pas sémites. Près
de 10% sont des Celtes et un incroyable 52% sont
un ADN-Y slave.
Les études concluent également à l'homogénéité
de l'haplotype modal R1a1 de sorte que l'ancêtre
a existé peut-être seulement 663 années
auparavant. La croissance peut avoir été
seulement d'une personne. Cela aurait coïncidé
avec la mise en place des communautés juives de
Mayence et de Worms (cf. Encycl. Judaica,
1972). Les motifs en forme d'étoile de l’ADN-Y
R1a1 indiquent une croissance rapide de la
population dans la population juive ashkénaze
dans les 1000 dernières années. La diversité
dans les Lévites Ashkénazes suggère une
intrusion génétique après la scission ashkénaze
d'autres populations juives (voir Behar et al.,
ibid., et Tableau 5). L'origine des Lévites
Ashkénazes R1a1 est donc probablement des
Khazars sorabophones convertis au Judaïsme et
entrés en Lévi et qui ont ensuite connu une
expansion rapide en Europe.
Parmi les tests effectués sur les populations
juives, nous pouvons comparer les Cohenim
séfarades et les Cohenim ashkénazes et nous
constatons que même dans les Cohenim il y a des
lignées hamites et japhétiques (Behar et al.,
ibid., Tableau 1). Alors que le Haplogroupe J a
une fréquence de .8684 pour les Cohenim
ashkénazes (AC) et .7536 pour les Cohenim
séfarades (SC), il y a toujours une fréquence de
sacrificateurs non-sémitiques entre 14,16 et
24,64 pour cent.
Il s'agit des Cohenim non-Aaroniques et même
non-Israélites :
Hamitiques
BR (ex DE JR) à .0263 pour AC et .0290 pour SC.
Il y a des E (ex E3a) à .0395 pour AC et .0435
pour SC.
Cohenim japhétiques présumés
K (xL, N3, O2b, P) à .0263 pour AC et .0290 pour
SC.
P (xR1a) .0263 pour AC et .0870 pour SC.
R1
a1 .0132 pour AC et .0580 pour SC.
Des populations juives de base seulement 37%
environ sont des Sémites réels dans les
échantillons de fréquence (Behar et al., Tableau
1).
Hg J est pour les Israélites ashkénazes (AI)
.3700 et pour les Israélites séfarades (SI), il
est .3651.
Les groupes hamites sont les suivants :
Y (xBR, A3b2) .0159 pour SI
BR (x DE JR) .1400 pour AI et .0794 pour SI
E (xE3a) .2200 pour AI et .1905 pour SI
Les lignées japhétiques présumées :
P (xR1a) .1500 pour AI et .2063 pour SI.
R1a1 .0400 pour AI et .0159 pour SI
À noter qu'il y a des pourcentages beaucoup plus
petits de Khazar convertis entre l’origine
généalogique de base israélite que parmi les
Lévites ashkénazes. Les conclusions qui peuvent
être tirées de ces statistiques c’est que les
familles de la noblesse, et peut-être des Kagan
et des Beck des Khazars, ont été converties et
ont reçu l'état Lévite, et/ou que les convertis
qui sont restés fidèles au Judaïsme sont allés à
l'ouest, en Europe avec les Huns, qui ont été
suivis par les Bulgares. Les Slaves paganisés
sont restés séparés ou derrière pour être
emportés par les Russ, puis les Mongols en 1215,
et sont venus plus tard en tant que les Sorbes
et les Belarus après les Huns et les Polonais.
Ce qui est certain, c'est que les partisans du
Judaïsme rabbinique ne sont pas entièrement
juifs ou israélites, et 63% de tous les Juifs
sont des convertis soit hamites ou japhétiques
au Judaïsme, c'est-à-dire, à moins qu'il puisse
être démontré que certains aspects de K sont
sémitiques plutôt que japhétiques, ce qui
projetterait le séquençage de l’ADN-Y en
révision complète.
Ce qui est intéressant dans les lignées K2 c’est
que parmi les 117 hommes de l’Haplogroupe K2
dans les banques de données de l'Université de
l'Arizona, près de 15% sont Juifs. L’ADN-Y K2
est originaire du Moyen-Orient. Le Dr Wells est
d'avis que l’ADN-Y K2 est phénicien antique.
Cela voudrait dire que les Phéniciens étaient
bibliquement de Japhet, mais nous savons
historiquement que ce sont des Édomites qui se
sont mélangés avec des Hamites. Ainsi, ils sont
sémites et hamites. Les seules nations
japhétiques commerçant en mer alliées avec eux
étaient les navires de Tarsis et des Kittim, les
Dodanim et ceux de Javan. Si K2 est en effet un
ADN-Y majeur phénicien, alors cela démontre que
les Anciens Rois de la Mer étaient une
agrégation des alliances commerciales des trois
subdivisions ethniques des fils de Noé. Cela
donc se reflète dans les conversions au Judaïsme
parmi les partenaires commerciaux.
Les Fils de Sem
Les fils de Sem sont Élam, Assur, Arpacschad,
Lud et Aram.
Nous
savons avec certitude que les Élamites sont
devenus les Perses, les Fils d'Assur sont
devenus les Assyriens, les Fils d’Arpacschad
sont devenus les Hébreux, et Aram est devenu les
Syriens et la source du nom de la langue
araméenne. Ils sont également allés plus tard,
vers le nord en Arménie.
De
l'échantillonnage de l'ADN moderne du projet
assyrien, les Assyriens modernes sont L, G, J et
certains R1b1, mais il semble que les R1b1
testés sont anglais du côté paternel et
assyriens du côté maternel. Ainsi, le Nord
d’Aram (dans le domaine de l'Arménie et la
Géorgie) et Assur ont développé le Haplogroupe
G, qui est l'endroit où il se trouve dans les
plus grands pourcentages.
Josephus
nous donne une indication claire des fils
d'Aram, quand il dit que sur les quatre fils
d'Aram, Uts a fondé Trachonite et Damas, qui est
le pays entre la Palestine et Ceolesyria, Hul a
fondé l’Arménie et Guéter les Bactriens, et
Masch les Mesaneans maintenant appelé Charax
Spasini. À partir de ce dossier, le Haplogroupe
G sémite semble venir d'Aram et Assur. Josephus
enregistre que Abraham s’est d'abord établi avec
une armée à Damas (ibid., Whiston tr., p. 32).
Josephus
dit que Jokthan des Hébreux avait treize fils :
Elmodad, Saleph, Asermoth, Jera, Adoram, Aizel,
Decla, Ébal, Abimael, Sabeus, Ophir, Eulat, et
Jobab. Il dit que ces personnes vivaient en Asie
de la Rivière indienne, Cophen, et les terres
qui l'entourent. C'est Kaboul moderne et dans la
vallée de sa rivière. Le terrain le plus vers
l'Indus est la Bactriane. Ainsi, si cela est le
fait de la question, il semble qu’ils se soient
déplacés à travers la péninsule arabique et la
plupart sont allés en Inde. Le résultat pourrait
être des groupes composites de Jokthan dans ce
qui est l'Afghanistan, Guéter à la Bactriane,
puis Lud dans le Pendjab. Le Haplogroupe
sémitique H en Inde serait alors dérivé d'une
mutation qui s'est produite chez les Hébreux de
Jokthan et les Sémites de Lud. Le coin d'or
d'Ophir se réfère aux terres de l'un des fils de
Jokthan qui s’est installé en Afghanistan ou en
Inde, et donc la zone d'Ophir peut être de
l'Hindu Kush en Inde ou peut-être, encore plus
probable, au Sri Lanka (Ceylan).
Afghans et autres Pashtounes
L’article de Wikipédia sur les Pachtounes
dit :
"Selon la plupart des anthropologues, les
Pachtounes de langue pashto semblent être
principalement d'origine iranienne (aussi bien
que d'être modifiés par diverses autres
envahisseurs et migrants au cours des siècles)
et sont très semblables aux Perses, aux Kurdes,
aux Tadjiks et aux Baloutches. Ces
anthropologues croient que les Pachtounes ont
des origines iraniennes orientales puisque la
langue pashto est classée comme une langue
orientale de l'Iran de loin lié à l'ossète parmi
d'autres langues iraniennes (voir Ethnologue
pour plus de détails).
Les Pachtounes Pashto-saxons se désignent
eux-mêmes comme les Pachtounes ou Pukhtuns selon
qu'ils sont des locuteurs du dialecte du sud ou
du dialecte du nord, respectivement. Ces
Pachtounes forment le noyau des Pachtounes
ethniques qui sont principalement un peuple
iranien et que l’on trouve dans le sud et l'est
de l'Afghanistan et l'ouest du Pakistan.
Beaucoup de Pachtounes de langue pashto se sont
cependant mêlés à différents envahisseurs,
groupes voisins, et migrants (comme l'ont fait
les autres peuples iraniens), y compris
peut-être les Ghilzai qui ont pu s’être mêlés à
des tribus turques, les Durrani qui ont interagi
considérablement avec les Tadjiks (un autre
peuple iranien), et les tribus pachtounes au
nord de Peshawar, qui se sont mêlées avec des
groupes dardes. En termes de phénotype, les
Pachtounes de langue pashto en général sont
principalement un peuple caucasoïde avec des
caractéristiques méditerranéennes, mais les
cheveux blonds et les yeux bleus et verts ne
sont pas rares, surtout parmi les tribus
montagnardes reculées."
Il dit aussi que :
"Dès le 1er siècle AEC au 5ème
siècle EC les régions où vivent les Pachtounes
ont vu d’immenses migrations de peuples de
l'Eurasie, y compris les Aryens, les Perses, les
Sakas, les Scythes, les Kouchans, les Huns, et
les Grecs. Plus tard, les envahisseurs
incluraient les Arabes musulmans et les tribus
turcophones d'Asie centrale ainsi que les
assauts dévastateurs des Mongols."
Il y aurait donc des ADN-Y R1a et R1b
japhétiques et C hamitiques présents.
L'article dit qu’à partir de :
"échantillonnage aléatoire de populations
pachtounes du Pakistan occidental) la preuve
anthropologique que les Pachtounes de langue
pashto sont un peuple indo-européen lié à
d'autres groupes iraniens ainsi que des
locuteurs de langues dardes comme le Kalasha et
le Nuristanis semble très probable, mais loin
d’être concluante en ce moment. Le test, bien
que toujours dans ses phases initiales, n'a pas
montré de lien substantiel entre la population
pachtoune générale échantillonnée avec des
marqueurs génétiques trouvés chez la plupart des
Grecs, des Juifs, ou des Arabes. Ce qui peut
être le cas c’est que les Pachtounes ont été
légèrement modifiés au fil du temps par divers
envahisseurs, tout en conservant leur base
orientale de l'Iran génétiquement globale.
Finalement, plus de recherche et un plus large
échantillonnage de l'ADN seront nécessaires
avant que des conclusions puissent être jugées
concluantes et généralement représentatives des
Pachtounes de langue pashto."
Les conclusions ici proviennent de la prémisse
que les Haplogroupes sémitiques sont tous J tels
que trouvés parmi les trois populations
mentionnées.
Les Pachtounes sont considérés comme iraniens,
qui, comme nous l'avons vu sont principalement
de le Haplogroupe I, mais ils varient en un
regroupement d’ADN-Y composite typique de l’Asie
centrale. Une tentative a été faite pour classer
les Pachtounes au 16ème siècle comme
l'un des Bani Israël, des Dix Tribus Perdues et
comme Joseph. Cela a été démystifié basé sur
leurs affinités indo-iraniennes et leur langue.
Cependant, c’est un fait historique que la zone
de Kandahar, Kaboul et la Bactriane aussi, de
l'autre côté de l’Hindu Cush, ont été faites des
provinces ou satrapies de l'Empire perse et
leurs langues sont donc affectées par ce fait.
Les similitudes de l'ADN avec Élam et leur
rapport historique de descendance et d’affinité
avec Jokthan indiquent que l'ADN original des
Sémites, y compris les Hébreux Jokthan, n'était
pas J, mais F et le dérivé primaire sémitique
est I et les dérivés secondaires sont J, et
ensuite H et G. Tous les Sémites de l'Est de
l'Élam et Jokthan étaient Hg I, plutôt que J.
Ainsi, ces mutations ont eu lieu au cours du
second millénaire AEC (avant l’ère courante) et
les modèles mathématiques étendus des
évolutionnistes sont faux.
Les
tribus Durrany et Galzay sont tenues pour être
les descendants directs des Ibrani ou Hébreux,
et probablement des fils de Jokthan. Certains
soutiennent également que les Rabbani, Shinwari,
Levani, Deftali, et Jaji de l'Afghanistan, et
les Efridi et Yusufzai en provenance du Pakistan
sont aussi des tribus hébraïques.
Lud
Lud est
une source d'incertitude et il y a deux
explications pour l'identification et les
emplacements des fils de Lud. Nous reviendrons
sur ce casse-tête sous peu.
Autres Emplacements des Hébreux
L'origine
et l'emplacement des Hébreux sont identifiés
avec Abraham comme venant d'Ur de Chaldée et les
fils d’Arpacschad sont identifiés avec les
groupes akkadiens dans la plaine mésopotamienne
entre les enfants d'Assur, ou les Assyriens, et
les fils d'Élam, qui étaient à l'Est du bassin
du Tigre sur le plateau persan.
L'autre
groupe des Mèdes, qui ont été associés avec les
Perses et ont occupé le haut pays, au nord de
l'Élam et Gutea, n’est pas sémite – ils sont les
fils de Madai, une tribu japhétique, et sont
identifiés aujourd’hui comme les Kurdes. Nous
allons y revenir plus tard. Les Guteans ou Guti
pourraient bien aussi être associés avec les
Goths comme éléments de la Horde parthe ou
scythe ultérieure.
Le Professeur Cyrus Gordon soutient également
que les Minoens primitifs sont des Sémites et
leur langage Linéaire A comme étant sémitique.
Son ouvrage sur Linéaire A a été reçu avec
controverse.
Les fils d’Arpacschad sont Schélach et Héber
(d'où le nom Hébreu est dérivé). Ses fils
Péleg et Jokthan ont vu la division dans les
Hébreux. Les fils de Jokthan étaient au nombre
de treize et ont occupé toute la zone orientale
de la péninsule arabique (Genèse 10:21-32), mais
beaucoup sont allés en Afghanistan ou au
Pakistan, comme nous le voyons ci-dessus. Les
fils d'Abraham à travers Agar et quelques-uns
des fils de Ketura, comme Madian, se joignirent
à eux plus tard. La lignée d'Abraham par Péleg
était de Rehu, Serug, Nachor, Térach à Abraham,
Nachor et Charan. Abraham a été appelé d'Ur en
Chaldée et envoyé dans le pays de Canaan.
Il s'ensuit que les lignées des pays sémitiques
nous donneront des indices quant à la
distribution des Haplogroupes et des mutations
sémites. La position de départ était de dire
simplement que le Haplogroupe J est le
Haplogroupe sémite et tous les autres ne sont
pas sémites, mais cela ne peut pas être vrai
étant donné la distribution et les mélanges des
groupes de nations.
Par exemple, dans la structure génétique des
Perses ou Élamites, Hg I est prédominant. Il
dépasse J d'environ 10%, et a un élément de F et
G avec elle. Ces groupes constituent la moitié
de l’ADN-Y des Perses. Les Arabes du
Moyen-Orient ont également une quantité
importante de I et certains G. Les Haplogroupes
G, I et J représentent la moitié des
Géorgiens/Arméniens et des Turcs, tandis que F,
G, I et J sont à moitié les Italiens avec I et J
comprenant la moitié de l’ADN-Y des Grecs. Quand
nous allons en Europe, l'équilibre de I augmente
de façon spectaculaire.
G,
(M201), H (M69), I (M170, M258, et P19) et J
(12f2.1) semblent être des lignées sémitiques
connues avec K (M9), la base de la racine de
tous les fils de Japhet. L’Hg J diminue à mesure
qu’il se déplace à l'ouest en Europe, mais l’Hg
I reste plus ou moins généreusement important à
l'ouest jusqu'aux Anglo-Saxons, puis diminue
parmi les Basques, les Gallois, les Irlandais et
les Écossais – mais il est néanmoins toujours
présent dans des quantités allant jusqu'à 15%
parmi ces Celtes connus.
Les Anglo-Saxons sont venus du Moyen-Orient
comme élément de la Horde à la chute de l'Empire
parthe de ce qui est maintenant la région de
l'Irak et la région au nord de celui-ci. Cela
s'est produit à la fin du deuxième siècle EC
(ère courante). Ils se sont déplacés dans la
zone du Nord-ouest de l'Europe et la Horde s’est
scindée en plusieurs vagues. Les Anglo-Saxons et
les Jutes et les tribus associées des Lombards,
des Danois, des Norvégiens, des Saxons et des
Fris, se sont divisés, mais ont conservé une
diversité similaire. Les Français du Nord sont
également du Moyen-Orient. Les Normands
constituent un élément et sont principalement
R1b mais ils contiennent d'autres groupes d'ADN
tels que I. Les autres sont des Francs en deux
groupes. Ce sont les Francs Riphatiens et les
Francs Saliens. Les Francs Saliens pouvaient
hériter à travers la lignée masculine seulement,
mais les Francs Riphatiens pouvaient hériter à
travers les deux lignées. Comme nous le savons,
Riphat était un fils de Japhet, et nous savons
que la noblesse de ces personnes a également
revendiqué être descendante de Antenor I, Roi
des Cimmériens, et aussi des Troyens. Ils ont
été interprétés comme la progéniture du groupe
qui est resté au Moyen-Orient avec les fils
d'Hector. Ils ont nommé leur grande ville
d’après Paris et Troyes. Beaucoup de ces
personnes se sont installées à travers la Manche
en Grande-Bretagne et ces tribus (par exemple
Parisii) étaient également des Celtes R1b. Plus
de 35% de la partie nord française sont des
Haplogroupes I et J. Environ 35% des
Anglo-Saxons sont Hg I du Moyen-Orient avec une
petite quantité de J. Environ 40% des Anglais
sont du Moyen-Orient et apparemment Sémites. La
majorité de tous les Celtes du Nord-Ouest et des
Anglo-Saxons sont R1b et une partie des mêmes
divisions génétiques que les tribus japhétiques
connues. L'exception évidente est à l'Est, où
les Slaves sont R1a, et les divisions plus
grandes R1a parmi la Horde à venir dans le
Nord-Ouest ont été parmi les Norvégiens, à
environ 30%.
Alliances d'Israël avec Japhet
Les travaux historiques des Britanniques nous
disent que les Gallois venaient du Moyen-Orient
après la chute de Troie. Ils ont rencontré les
Cornouaillais en Afrique du Nord, qui étaient
aussi des survivants de la chute de Troie. Nous
savons maintenant que c'était le royaume troyen
de Wilusia. Leurs histoires disent qu'ils ont
trouvé les fils de Magog déjà en Grande-Bretagne
et les ont subjugués. Ils étaient les fils de
Riphat, et au Moyen-Orient ils faisaient partie
de l'empire hittite, en alliance avec les Hatti
et Kalti plus à l'est.
Les Gallois sont connus comme le peuple Kymry et
ont autrefois occupé toute la Grande-Bretagne et
une partie de la France comme et avec les
Gaulois. Ils ont été confinés à l’Armorique, qui
est maintenant la Bretagne. Ils étaient tous
Celtes (Gimirra de Gomer). Ils sont à
l'origine des Cimmériens (c.-à-d. Gomeriens) qui
ont ré-envahi l'Anatolie et établi la Galatie.
Ils formaient la garde de Cléopâtre de 400
hommes que César a présenté à Hérode. Ils sont
probablement à l'origine des R1b en Turquie à
>15% et dans une moindre mesure en Syrie. Les
Turcs font aussi partie du groupe linguistique
ouralo-altaïque et donc certains peuvent être
R1a et R1b de cette source également. Ces deux
groupes et Q sont très proches et la division
entre les groupes est sautée par une très petite
mutation. Ainsi, les Indiens d'Amérique sont des
parents éloignés des Celtes et des Slaves.
Irlande
Les
premiers habitants de l'Irlande étaient les
Nemediens et ensuite les Formoriens, les
Firbolgs (ou les gens en cuir) et les
Tuatha
de Danaan.
Les premiers descendants sont venus de l'Espagne
et ont été inscrits comme Gadeliens, les
descendants de Gaodhal, sixième de Japhet (op.
cit.). Les Gadeliens étaient des Scythes qui se
sont étendus des Boristhenes aux Massagetae (la
Grande Horde Gothique) et des Saces (de là
Scythes) à la Mer Caspienne. Ils étaient des
nomades
(op. cit. p. 61).
Génétiquement, nous pouvons les identifier comme
les fils de Gomer et de Magog. Les variations de
l’ADN-Y montrent que les Juifs ashkénazes sont
plus R1a1 que R1b, et de là, les variations
doivent être arrivées après les divisions des
groupes tribaux. 52 % de tous les Lévites
ashkénazes sont des fils de Gomer. Ainsi les
prophéties quant à Gomer ont la pertinence
directe vers Israël aujourd'hui.
Une
variété ancienne d’ADN-Y RxR1 est trouvée en
Afrique au Cameroun. Elle peut s’être produite
avec les mouvements tribaux des tribus
japhétiques en Afrique, soit de l'Est quand les
groupes Aborigènes australiens RxR1 sont partis
pour l'Australie suivant les nomades Cuschites
Hg C avant eux, ou même aussi tard que les
Vandales qui ont envahi l'Afrique du Nord de
l'Espagne et ont été après vaincus.
Nous savons par l'histoire irlandaise et les
généalogies que les Milésiens étaient des fils
de Magog, et ils ont été testés à R1b.
Cependant, nous savons qu'ils sont entrés en
Irlande un peu tard et ont remplacé une tribu
qui était là auparavant. Cette tribu a régné en
Irlande pendant des centaines d'années et était
connue comme les Tuatha de Danaan, appelée la
tribu de Dan ou les Danaens. Ils sont tenus
d'avoir aussi navigué avec les Philistins, les
premiers Grecs et apparemment les Phéniciens (Danaans
and Danites – Were the Hebrews Greek?
BAR,
Juin 1976). Il est fort probable que cette tribu
soit responsable de l'incidence de l'Haplogroupe
I en Irlande puisque les Anglo-Saxons n'ont pas
occupé l'Irlande quand ils sont entrés dans la
Grande-Bretagne, tandis que les Normands sont
majoritairement R1b avec quelques I à partir des
Anglo-Saxons, mais le pourcentage I ne
représenterait pas proportionnellement le
pourcentage en Irlande. La proportion de I est
presque aussi élevée que la proportion de I en
Écosse, mais l’ADN-Y prédominant là aussi est
R1b. Il peut y avoir une infusion pourcentage de
I de la Norvège dans les deux zones. Si le récit
historique et son interprétation selon laquelle
les Tuatha de Danaan sont en effet une branche
de Dan d'Israël, alors nous avons le lien
démontrable qui montre les tribus du Nord comme
Haplogroupe I. Ainsi, 40% des Anglo-Saxons
seraient Sémites et probablement d’Israël, comme
les registres l’affirment. L'argument contre cet
aspect, c'est que, des Juifs, moins de 2% sont
de l’Haplogroupe I. Lorsque l'on considère,
cependant, que seulement 37% des Juifs sont des
Sémites, cela apporte l'échantillon jusqu'à
environ 6% des Juifs connus et donc peut bien
représenter le reste des autres tribus qui sont
restées avec ou sont retournées auprès de Juda.
Il y a aussi une incidence connue du K2
phénicien à partir des navires de Tarsis qui
sont présents à Tyr, à Malte et au Pays de
Galles. Thomas Jefferson, le président américain
a revendiqué descendre du Pays de Galles et
était de ce sous-clade gallois de l'Haplogroupe
K2.
La
première colonie en Irlande a été commandée par
Partholan, qui était de la lignée ancienne des
Grecs avant que la terre de la Grèce n'ait été
envahie et peuplée par les fils E3b de Cham de
l'Afrique. La deuxième colonie était commandée
par Nemedius, qui s'est bien battu pour
consolider sa colonie. Cependant, les Formoriens
ont envahi et ont vaincu Nemedius dans Cork et
ont repris l'Irlande (MacGeoghegan and Mitchell,
History
of Ireland, 1867, p. 54).
Les Firbolgs sont identifiés par MacGeoghegan
comme étant les Belges et il y avait une tribu
de Belgae en Grande-Bretagne. Les Formoriens ont
été aussi vaincus, ou ont abandonné la terre et
les Firbolgs en ont pris possession. Ils ont été
remplacés par les Tuatha de Danaan, les
descendants de Nemedius qui sont retournés en
Irlande par l'Allemagne et via la Norvège et le
Danemark (MacGeoghegan ibid. p. 55).
MacGeoghegan prétend qu'ils peuvent avoir été
des descendants des Dumnoniae de Cornwall.
Il
déclare aussi qu'ils peuvent avoir été des
descendants de Jobath, petit-fils de Nemedius,
qui a mené une partie de la colonie en Allemagne
du Nord de l'Irlande à l'origine (ibid. p. 55).
Ainsi, les colons originaux de l'Irlande étaient
une alliance de Grecs antiques et des Tuatha de
Danaan ou la Tribu des Danites, qui étaient des
fils de Nemedius. Ceux-ci étaient les premiers
Danites navigateurs alliés avec les Rois Anciens
de la Mer.
MacGeoghegan affirme également que Briotan Maul,
le petit-fils de Nemedius par Feargus, s'est
établi avec sa tribu en Grande-Bretagne, qui,
selon le
Psautier de Cashel, a été appelée selon son
nom – et d’après certaines informations, ils se
sont installés là comme des Bretons.
MacGeoghegan dit que c'est au moins aussi
crédible que le compte rendu par Geoffroy de
Monmouth. Cela représenterait le Haplogroupe I
(sous-clade des Îles) parmi la distribution des
Pictes R1b en Écosse. Ils ne sont pas venus de
la Norvège et sont presque exclusifs aux Îles
Britanniques. Les Pictes ont aussi été des
premiers colons en Grande-Bretagne pratiquement
des millénaires avant les Écossais.
En même temps, soit par la défaite ou le départ
des Formoriens, un autre groupe de 5000 hommes
est entré et a divisé l'Irlande dans une
Pentarchie avec le gouverneur Slaingey de
Leinster chef de la Pentarchie de Slaingey, Rory
(Rughruighe), Gan, Gannan et Sengan – tous les
enfants de Dela de la race des Nemediens. Les
gens étaient alors connus sous trois noms
différents : Galleniens, Damnoniens et Belges.
Ils ont régné pendant quatre-vingts ans sous
neuf rois, le dernier était Eogha. Il a épousé
la fille d'un prince espagnol. Elle a été
enterrée à Tailton dans Meath, qui a été nommé
d’après elle.
Les Belges ou Belgiae ont été localisés les plus
proches des Dumnoniens en Angleterre aussi et il
est probable que les trois noms étaient
indicatifs d'une alliance des trois tribus qui
se sont déplacées de l'Europe à l'Angleterre et
en Irlande. Les Galleniens étaient apparemment
des Gaulois, de là aussi Galates. Cependant, il
est probable que par les Nemediens sous les
groupes des Tuatha de Danaan que nous voyons le
Haplogroupe I s’imposer avec le R1b (AMH) des
Celtes. Ce groupe est venu du Danemark beaucoup
plus tôt que le R1b postérieur des Danois de la
Horde Parthe (les Danois postérieurs étaient DYS
390/391 23/11 et il y a deux groupes I au
Danemark aussi qui diffèrent, un étant le type
Anglo-Saxon, voir ci-dessous). Le mouvement
pourrait aussi avoir été avec les Huns ou même
avant eux des groupes de la navigation dans le
deuxième millénaire AEC. Il y avait de l’ambre
et des systèmes de commerce étendus dans la
Baltique à ce moment-là. Comme on le voit
ci-dessus, nous savons que les commerçants K2
Tarsis/phéniciens étaient au pays de Galles et à
Cornouailles et cette lignée existe toujours là.
C’est à
partir de ce moment-là que les Tuatha de Danaan
sont descendus en Irlande sous Nuagha Airiodlamh
qui a vaincu les Firbolgs sous Eogha dans le
Comté de Mayo. Les Firbolgs ont été chassés et
ont cherché le refuge dans les îles du Nord
(ibid. p. 55). Ces gens qui se sont installés
dans les Îles étaient R1b. Aussi, comme nous
avons noté, il y a le sous-clade de I1b2a−Îles
et ce haplogroupe est trouvé presque
exclusivement dans les Îles Britanniques et
fortement en Écosse. Ce n'est pas le Viking
norvégien. Le même Haplogroupe I est trouvé
parmi les Basques. Il apparaît qu'il n'est
certainement pas venu de la Scandinavie, mais
plutôt des Tuatha de Danaan venus du
Moyen-Orient en Espagne et certains sont restés
parmi les Basques. L'élément principal a
continué en Irlande et est allé en Écosse et
dans les Îles. Ils sont trouvés parmi les Pictes
aussi, donc ils doivent être partis de là avant
les Écossais qui se sont déplacés de l'Irlande à
Alba, puisque l'Écosse a été nommée avant que
Niall des Neuf Otages ne l'ait renommée. Ils ne
sont pas venus de l'Europe avec les Écossais ou
les Milésiens. La signification de la Tribu de
Dan comme les Tuatha de Danaan est que ce n’est
pas limité à l’Irlande comme plusieurs des
Israélites britanniques ont pensé. Ce groupe est
concentré en Écosse aussi avec des nombres plus
petits dans les Gallois, avec certains en
Angleterre et peut-être parmi les Basques. Du
chapitre 7 d’Apocalypse, nous voyons que Dan et
Éphraïm cessent d'exister comme des entités
séparées et le composé Joseph émerge de nouveau,
mais cette fois sans Manassé qui est séparé.
Les Îles
sont celles des côtes de l'Écosse puisque ce
n'était pas avant quelques siècles que quelque
colonisation que ce soit de Thule ou de
l'Islande n’ait été faite.
Les
Milésiens sont venus via l'Espagne en Irlande et
une sous tribu est entrée en Angleterre de
l'Espagne et sont devenus connus comme les
Brigantes. Ils se sont établis à l'ouest de
l'Angleterre dans ce qui est maintenant
approximativement le secteur de Lancashire.
Les
Irlandais, jusqu'aux Milésiens entrés vers 500
AEC comme nous le voyons ci-dessus, ont été
compris de trois groupes appelés les Formoriens,
les Firbolgs et les Tuatha de Danaan ou la Tribu
des Danites.
Les
Milésiens que nous connaissons à coup sûr sont
des Celtes R1b et nous avons leurs généalogies
et leur ADN pour correspondre à ces généalogies.
Ces Celtes s’harmonisent avec les autres
séquences celtes ADN-Y R1b, mais diffèrent selon
les régions et les tribus.
Le
facteur principal déterminant dans les
Haplogroupes celtes R1b à l'Ouest de l'Europe
est trouvé dans les divisions de l’Haplogroupe
R1b1. Quatre sous-clades de R1b sont dérivés des
répétitions des marqueurs DYS 390 et 391.
Le Haplotype Modal Atlantique (AMH), qui a une
valeur de 24/11 pour 390/391, est trouvé sur la
côte atlantique de l'Espagne à l'Écosse. Ces
Celtes sont ces vieux habitants de l'Europe et
descendent de Japhet par les divisions
précédentes et sont trouvés parmi les Basques.
Les
Anglo-Saxons sont venus en Grande-Bretagne après
l'occupation romaine en Grande-Bretagne en 410
EC et avant la Conquête Normande. Les
Anglo-Saxons ont les deux Haplogroupes R1b et I.
Le sous-clade R1b (R1b1) des Anglo-Saxons est
distingué par les marqueurs DYS 390 et 391 qui
ont les valeurs de 23/11. Ces valeurs sont
trouvées au Nord-Ouest de l’Europe parmi les
autres nations de la Horde qui s'est établie là
en Flandre (la Belgique), aux Pays-Bas, au
nord-ouest de l'Allemagne et au Danemark. Les
Fris et les Anglo-Saxons partagent ce marqueur
d'identification parmi les autres. Eux et les
Saxons de l'Allemagne sont pratiquement les
mêmes gens.
Les
autres sous-clades de R1b sont des répétitions
de 24/10, qui sont trouvées partout en Europe et
les répétitions de 23/10 qui sont trouvées au
sud-est de l’Allemagne et en Autriche. Ainsi,
ils sont tous de différentes lignées celtes ou
fils de Gomer qui sont venus à l'origine de la
Scythie et la Parthie.
Les
variantes de l’Haplogroupe I qui sont trouvées
parmi les Anglo-Saxons de I1a sont I1a AS (1-8).
I1a AS est la variante la plus commune de
l’Haplogroupe I1 et est nommé parce qu'il est
trouvé où les Anglo-Saxons à l'origine se sont
installés dans leur mouvement du Moyen-Orient.
Cela tombe comme nous nous déplaçons dans le sud
de l'Allemagne de l'Est et au-delà. Il est
probable que les variantes de ce Haplogroupe
marquent les déterminations héréditaires de
certaines des Dix Tribus. Ces nations sont
invariablement liées avec les Celtes hittites
comme c’est décrit ci-dessus. L'AMH est un
groupe précédent distinct des Anglo-Saxons qui
avaient des marqueurs distinctifs tant sur R1b1
que sur I1a AS.
La
détermination des Îles I1a AS quant aux Danois
par opposition aux comptes rendus des ancêtres
allemands sur le continent rapportera plus
d'informations. Le I1a- (Nordique) est la forme
la plus populeuse trouvée en Suède et au
Danemark et est la deuxième en Norvège. Il est
trouvé dans seulement des petites quantités en
Europe Continentale au sud des Mers du Nord et
la Baltique et prend la deuxième place à I1a –AS
au Danemark. Ainsi les groupes I se mélangent au
Danemark mais les Danois sont principalement des
types Anglo-Saxons (voir le document
Population Varieties within Y-Haplogroup I and
their Extended Modal Haplotypes (and also R1b
Varieties) par Ken Nordtvedt sur
www.northwestanalysis.net)
I1a uN
(ultra Nordique) atteint son sommet en Norvège.
Il diffère du Nordique I1a par un changement de
DYS 385 b de 14 à 15. Il se divise aussi en deux
variétés basées sur la valeur répétée DYS 461.
I1a uN1 a des modaux 12, 28 à DYS 461 et 449, ce
qui est commun à la plupart des I1a. I1a uN2 a
11, 29 répétitions à DYS 461 et 449 et sont plus
fortement associées à la Norvège qu’à la Suède
et au Danemark. Ces divisions peuvent s’être
produites après la Grande période de Migration
en Scandinavie du troisième et quatrième siècle
EC.
I1a1 (vieux I1a4), est un petit sous-clade de
I1a trouvé dans environ 10 % des I1a provenant
de l'Europe de l'Est.
I1b1 Din (Dinarique) prend son nom d'une chaîne
de montagnes dans les Balkans près de là où
c'est le plus prolifique. Il s'est aussi étendu
à travers la plupart de l'Europe de l'Est.
I1b1 –Ouest (Occidental) est une variété de
(vieux) I1b, P37.2 + est trouvé dans des
quantités prolifiques à travers des secteurs
côtiers de la Mer du Nord et la Baltique de
l'Allemagne et s’étend ensuite dans les Îles
Britanniques. Il est notamment identifié par 15
répétitions DYS 388 au lieu des 13 répétitions
habituelles de Dinarique I1b1. Cette variété a
été aussi récemment trouvée en Estonie.
I1b1-Îles est une variété récemment découverte
de I1b1, qui semble résider principalement dans
les Îles Britanniques. Les marqueurs
d'identification sont DYS 385, 463 et 464.
I1b1a (vieux) I1b2, un sous-clade de I1b1 M26+
est un grand pourcentage de la population de
Sardaigne et s'étend dans l'Ibérie. Il n'est
trouvé dans aucun niveau au Nord de l'Europe.
I1b2a - Cont (Continental) (vieux I1c) est la
variété principale de I1b2a. C'est le plus dense
au nord-ouest de l’Allemagne, au Pays-Bas et
jusqu’au Danemark et un peu dans le sud de la
Suède et de la Norvège.
Une
quantité significative est trouvée aussi dans
les Îles Britanniques venant des
immigrants/envahisseurs germaniques et
scandinaves.
I1b2a-Root est une variété peu commune de I1b2a
qui s'étend de l'Italie et de l'Ibérie jusqu’au
Danemark et se scinde en deux variétés basées
sur quatre des marqueurs.
I1b2* est une autre variété qui est trouvée de
l'Italie et de l'Ibérie jusqu’au Danemark et se
scinde aussi en deux variétés basées
principalement sur le Marqueur DYS 448.
I1b* (vieux I1(y) est une petite variété de
l'arbre et est 10/10 DYS 459a,b comme une de ses
fonctions modales et 19.19 ou 19.21 à YCA11a,b
et 12,29 à DYS389I,ii.
Il est considéré que les variétés de ces
sous-clades de l’Haplogroupe I détermineront le
peuple hébreu et les divisions R1a et R1b
détermineront les branches de l'alliance hittite
formée avec la Scythie et la Parthie et les
Steppes progressivement en Europe.
Il est maintenant soutenu et accepté que les
Haplogroupes I et J sont des sous-groupes de F
qui était autrefois un groupe combiné IJ, étant
un seul peuple.
La Colonisation de l'Europe à partir du
Moyen-Orient et de la Scythie
Israël est entré en captivité sous les Assyriens
vers 720 AEC et a été amené dans le Nord dans ce
qui est devenu la Parthie et la Scythie. Là, ils
ont forgé une union avec le peuple celtique qui
formait les Hittites du Nord.
Il y avait des membres de la horde hittite et
scythe qui se sont déplacés après la chute de
Troie en Europe et en Grande-Bretagne et en
Irlande et ailleurs en Europe.
Sous Odin et après leur défaite par les Persans
en Parthie au deuxième siècle de l'ère courante,
les Parthes et une partie de la horde scythe se
sont déplacés au Nord-Ouest de l'Europe et ont
établi un certain nombre de nations incluant les
Anglo-Saxons, les Jutes, les Fris et les
Hollandais, les Saxons de l'Allemagne, les
Danois, les Norvégiens et certains des Suédois,
les Lombards, les Goths, les Suèves, les
Dalmatiens, les Vandales et d'autres. Ils ont
établi les nations d'Europe occidentale, de
l'Espagne et du Portugal au Danemark et en
Norvège à travers la Grande-Bretagne et Thule
(l'Islande) et le Groenland.
Les Slaves devaient se déplacer en dernier et
ainsi la division est de l'Europe occidentale
sous les fils R1b de Japhet et l'Europe
orientale sous les fils R1a de Japhet qui sont
tous mélangés avec les Sémites hébreux et les
fils d'Assur.
Les
Slaves de Rosh, Meshech et Tubal sont tous de
R1a, R1a1, les Slaves et les Scandinaves. Le
royaume russe a été établi par les Russ
Varangian de la Scandinavie qui ont établi des
royaumes basés sur Kiev et Novgorod de 850-950
EC.
De la Scythie à l'Irlande
Les Nemediens sont les ancêtres des Tuatha de
Danaan. La chronologie de l'occupation de
l'Irlande est intéressante.
MacGeoghegan, rapportant les histoires antiques,
dit que Fenius Farsa, le roi des Scythes avait
deux fils, Nenual étant l'aîné et l'héritier de
la couronne.
Le deuxième était Niul qui était devenu érudit
dans les langues multipliées à partir de la
confusion à la tour de Babel. Niul a fait un
voyage en Égypte où il a épousé Scota la fille
du Pharaon (Cincris) de l'Égypte et s'est établi
à la terre de Capacirunt aux frontières de la
Mer Rouge. Niul a eu, par Scota, un fils
Gaodhal. Il a été prétendument mordu par un
serpent et présenté à Moïse au moment de
l'Exode. La morsure est devenue verte et la
marque l’a fait mériter le nom de Gaodhal-Glas
ou Gadelas (glas
a la signification de
vert
dans la langue écossaise ainsi nommée de Scota
sa mère).
Ils faisaient ainsi partie de la
multitude
mélangée dans l’Exode. Les généalogies
milésiennes semblent faire cette revendication
aussi.
Ils se sont installés sur le secteur de l'Égypte
répartie à eux par le Pharaon pour la
progéniture de sa fille. Après trois
générations, ils ont été attaqués par les
Égyptiens sous Pharaon-En-Tuir et ont été ainsi
forcés de se déplacer, sous leur dirigeant Sur,
le fils d'Easur, le fils de Gaodhal, en Crète où
Sur est mort et a été succédé par son fils
Eibher, connu autrement comme Heber-Scot
(MacGeoghegan ibid., pp. 56-57).
Heber-Scot s'est déplacé de là à la Rivière
Tanais en Scythie via la mer Égée et la mer
Euxin. Ils ont vécu là pendant quatre
générations parmi les Scythes sous Agnamon,
Tait, Adnoin et Lamphion.
Les Scythes se sont étendus aussi loin que la
région autonome Uigur de ce qu'est maintenant la
Chine dans le deuxième millénaire AEC. Nous
avons trouvé leurs momies et des tissus kilts
écossais, très semblables aux tissus de chasse
écossais, bien préservés.
Les Gadeliens ont été soupçonnés et persécutés
et ont cherché refuge parmi les Amazones sous
Adnoin et sont partis sous son fils Lamphion
pour Gaethluighe. Certains pensent que c’est
Gothia, mais selon O’Flaherty c'est Getulia en
Afrique, où ils sont restés pendant huit
générations sous huit rois, étant les
descendants de Lamphion ; le dernier étant Bille
le père de Gallamh ou Mileag-Espaine (en latin)
appelé Milesius, qui était l'ancêtre des
Milésiens ou des Irlandais antiques. Nous savons
maintenant que les Amazones étaient des femmes
mariées des tribus scythes qui se sont battues
avec leurs hommes à cheval.
Milesius
est retourné pour une courte période en Scythie
où il a été reçu par Riffloir, le roi descendu
de Fenius-Farsa du fils aîné Nunnual. Il l'a
nommé général et lui a donné sa fille Seaug en
mariage par qui il eut deux fils, Donn et
Aireach surnommé Feabhrua. À la mort de sa
femme, et s’étant fâché avec son beau-père, il
est parti avec les garçons et sa troupe de
Gadeliens pour l'Égypte où il a reçu le
commandement de la force engagée dans la guerre
avec les Éthiopiens.
On lui a donné aussi une femme de Pharaon
Nectonibus, aussi nommée Scota, par qui il eut
deux fils, Heber-Fionn et Amhergin.
Dans ce temps, il avait douze jeunes hommes
instruits dans les arts et les sciences pour le
retour en Espagne.
Après sept ans en Égypte, il retourna en
Espagne. Après une grande sécheresse de
plusieurs années, les Gadeliens partirent pour
l'Irlande.
Ainsi, nous voyons les liaisons et les
connexions entre les Scythes et les Irlandais et
les Écossais. Les langues de P et Q Brittoniques
sont différentes mais elles sont historiquement
rapprochées des personnalités bibliques de Gomer
et Magog.
Étant donné la tranche de temps, nous sommes
aussi capables d'identifier une autre liaison
d’ADN-Y qui semble être les Nemediens ou Tuatha
de Danaan. Les Irlandais milésiens ont une
structure d’ADN-Y de base de R1b, qui est
commune aux Celtes. Elle a aussi un Haplogroupe
ADN-Y qui est I et semble être d'origine
sémitique.
Ce même Haplogroupe est aussi présent dans les
Anglo-Saxons et les Européens généralement dans
les niveaux de plus de 40 % comme nous le voyons
dans les Croates et les Norvégiens aussi. Ces
deux nations ont cependant un Haplogroupe slave
principal de R1a plutôt que R1b celtique, qui
est néanmoins présent dans les deux groupes.
En regardant les histoires et la structure de
l'ADN, nous sommes capables d’assembler le
mouvement des tribus et faire les connexions qui
seraient autrement impossibles.
Comment cet Hg I sémitique entrerait-il dans les
Européens ?
La réponse peut être trouvée dans les pages de
la Bible.
Il y
avait les fils de Ketura qui se sont déplacés
dans le Nord avant l'Exode et qui sont aussi
devenus les Grecs Spartiates comme nous le
savons de Josèphe (Josephus). Ces gens étaient
les gens qui se sont déplacés en Irlande et une
partie de ce groupe peut avoir formé les groupes
des Nemediens ou des Tuatha de Danaan. On pense
que les Tuatha de Danaan ont été associés avec
les Grecs et il est possible que les Danites
soient associés aux fils de Ketura et de là ils
expliquent le Haplogroupe I et aussi un peu du
Haplogroupe J des Grecs. L'E3b est une intrusion
postérieure des Cananéens phéniciens et de
l'Afrique.
Ces gens
sont aussi venus en vagues du Moyen-Orient. Les
Croates sont aussi très élevés dans le
Haplogroupe I et aussi élevés que les
Scandinaves en pourcentages. Ainsi partout dans
les Balkans, nous avons une population élevée de
l’Hg I de sources différentes, mais ayant
beaucoup en commun de façon ancestrale.
L'Alliance Hittite
Tôt dans la carrière du Roi David, il a établi
des alliances avec les royaumes du nord de Tyr
et Sidon et aussi avec les Hittites qui étaient
tant des fils de Riphath que des fils de Magog.
Ils ont été affaiblis par la perte du royaume
Wilusien dont la capitale était Troie. Ainsi les
fils d'Ashkenaz ont été exposés en Israël à ce
moment-là et les Israélites les ont rejoints et
les ont convertis plus tard.
Tant les fils d'Ashkenaz que les fils de Riphath
et les fils de Togarmah sont tous les fils de
Gomer. Néanmoins, Magog a aussi R1b. De là les
divisions de R1a et R1b sont arrivées après que
les tribus se soient divisées. Certains sont
devenus R1a et certains R1b. Les fils d'Ashkenaz
sont devenus R1a ou bien les groupes R1a se sont
appelés les Ashkénazes bien qu'ils soient
Slaves, ce qui est très possible suite à leur
exposition khazar.
Saül a consolidé le secteur d'Israël, mais David
l'a étendu à l’Euphrate.
Il était en Cappadoce et au nord de là en
Scythie parmi Méschec et Tubal au début du
onzième siècle AEC. Nous le savons des Psaumes
(Ps. 120:5).
Or, il y avait deux Méschec dans la Bible. Un
était fils de Japhet, l'autre était le fils
d'Aram, fils de Sem, vivant en Syrie. Il est
possible que David fût en Syrie parmi les fils
de Méschec, fils d'Aram, qui avait été identifié
avec les fils de Kédar.
Le composé Méschec et Tubal identifie les fils
de Japhet et non pas le fils d'Aram.
Le secteur de Cappadoce en Turquie qui a été
repris par Méschec il y a des millénaires a été
à l'origine appelé Kus d’après Cusch, le fils de
Cham et père de Nimrod. Le roi de l'empire
hittite avait sa capitale là et les Babyloniens
l'ont mentionné comme le Roi de Kus.
L'empire hittite a été divisé en des divisions
du Nord et du Sud. Elles étaient composées des
fils de Cham et des fils de Japhet les deux de
Gomer et Magog et Madaï ou les Mèdes, et les
alliances commerciales avec eux basées sur et
avec Sidon, de Tarsis et également les fils de
Heth. Homer, dans l’Iliade, dit que
l'armée troyenne parlait beaucoup de langues.
Les habitants de Tyr et du Liban devaient
devenir les descendants ADN-Y K2 des alliances
de la mer des fils de Japhet (fils de Javan
Tarsis et Elischa, et Kittim et Dodanim ou
Rhodanim). Ceux-ci n'étaient pas des fils de
Heth et nous les voyons dans les descendants du
Liban et de la Malte aujourd'hui.
Le Haplogroupe H
Le Haplogroupe H est largement confiné à l'Inde
au nord comme au sud, et à un moindre degré au
nord des Ouzbeks et Kirghizes. La plus grande
incidence dans le sud indique qu'il était là
avant l'invasion aryenne. S'il s'agit d'un
Haplogroupe sémitique, nous devons identifier la
nation sémite qui a muté dans ce Haplogroupe
dans l'isolement en Inde.
La tribu sémitique qui serait admissible est la
tribu des fils de Lud. On ne doit pas les
confondre avec les Ludim qui sont des fils de
Mitsraïm ou d'Égypte. Josèphe identifie ces
personnes avec les Lydiens qui occupaient la
région de la côte ionienne en Anatolie ou
Turquie moderne. C'est logique car ils étaient
alors à l'ouest d’Aram et au sud d'Assur et
d’Arpacschad, parmi les tribus sémites et au
Nord des Hamites. On en sait plus sur cette
section, mais eux et leurs descendants ne sont
pas Hg H.
Il y avait une tribu appelée les Lullubi dans la
région nord-est de ce qui est aujourd'hui l'Irak
qui a fait la guerre aux Akkadiens. On pense
qu’ils sont un clan de ce que sont aujourd'hui
les Kurdes appelés les Lullu, mais ce ne serait
pas les mêmes personnes puisque le corps des
Kurdes sont des fils de Madaï de Japhet ou les
Mèdes
–
mais ils peuvent très bien avoir été absorbés au
cours de cette période de temps.
D'autres, cependant, ont identifié les fils de
Lud comme se déplaçant à l’est de l'Élam. Cela
pourrait les mettre dans le domaine de l'Indus.
La cité antique dans le Pendjab est Ludhiana,
apparemment nommée d'après Lud. L'explication
serait donc que la majorité s’est divisée et est
allée à l'est et a établi les villes de l'Inde.
Cela expliquerait aussi pourquoi les anciennes
villes d’Harappa et Mohenjo Daro dans le bassin
de l'Indus ont eu des contacts avec les
Assyriens ca. 1750 avant notre ère et ont adopté
leurs systèmes d'adoration. Les Haplogroupes C
que l’on s'attendrait à trouver là-bas avec les
fils de Cusch sont compensés par les autres
groupes H, L et J dans le sud avec les plus
grands éléments aryens en Inde du Nord de R1a,
avec le plus ancien RxR1 des Dravidiens en plus
grand nombre que R1a dans le sud. L'Inde est
donc une combinaison des trois lignées, avec les
fils de Cusch se déplaçant vers le nord et
l'est.
Les Fils d'Abraham
Abraham a eu un certain nombre de fils et aussi
les tribus de Lot, son neveu, ont été associées
à lui. Lot a fondé Moab et Ammon dans ce qui est
aujourd'hui la Jordanie. Isaac a eu deux fils,
Jacob et Ésaü, et les fils d'Ésaü ont fondé Édom
et sont devenus les Édomites ou Iduméens qui ont
pris part au système commercial avec les
Phéniciens. Plus tard, ils se convertirent au
Judaïsme par Jean Hyrcan à l'époque des
Maccabées après 160 AEC. Ils ont été absorbés
dans la Judée et sont devenus les Juifs du sud,
par opposition aux Galiléens.
Jacob eut les douze tribus : Joseph et Benjamin
par Rachel, Dan et Nephtali par Bilha, la
servante de Rachel, Ruben, Siméon, Lévi, Juda,
Issacar sont nés à Léa, et Gad et Aser sont nés
de Zilpa, servante de Léa. Nous savons que des
fils de Léa, les fils de Lévi se sont développés
dans la lignée J2 (sur 1700 ans).
Cependant, pour que l'élément irlandais des
Tuatha de Danaan soit de la tribu de Dan, ils
devraient être Hg I. (Une autre explication
pourrait être qu’ils ont été complètement
éradiqués et les éléments Hg I sont de la
Norvège et l'Angleterre. L'objection à ce point
de vue c’est que le pourcentage de R1a devrait
être aussi grand qu'il est en Écosse, mais il
est minuscule en comparaison. Ainsi, le Hg I là
n'est pas de la Scandinavie ou de l’Angleterre
et n'est pas milésien non plus et est donc
Tuatha de Danaan.) La plupart des autres fils
d'Israël aussi devraient être Hg I aussi, de
sorte que les mutations auraient dû se produire
non seulement parmi les femmes, mais aussi entre
les fils des mêmes mères. L'autre explication
est que les mutations se sont produites en
Égypte et les tribus ont divergé là. Les mêmes
divergences ont dû se produire chez les Hébreux
Jokthan et les Élamites et les fils de Ketura.
À partir des proportions de l'ADN chez les
Élamites et les Arabes, il semble que l'ADN
précédemment se soit développé indépendamment
parmi les fils de F en I comme le Haplogroupe
majoritaire, mais aussi en J et G.
Il est enregistré [cf. ci-dessous] que les fils
de Ketura étaient Zimran, Jokschan, Medan,
Madian, Jischbak et Schuach. Les fils de
Jokschan étaient Séba et Dedan. Les fils de
Dedan furent les Aschurim, les Letuschim et les
Leummim. Les fils de Madian furent Épha Épher,
Hanoch, Abidah et Eldaa.
Nous savons que les Madianites ont été soumis à
la guerre en premier lieu dans les jours avant
qu'Israël aille en captivité en Égypte et plus
tard après l'Exode à cause de l'idolâtrie. Ils
devaient occuper la zone de l'Arabie avec les
fils d'Ismaël et les Hébreux Jokthan avant eux.
Nous savons d'après l'autorité de Josephus que
les Spartiates étaient les fils de Ketura.
Ainsi, la grande quantité de l’Hg I de l’ADN-Y
en Grèce peut être des Fils de Ketura, avec le J
venant plus tard à partir de la conquête de
l'Asie Mineure et des Arabes dans les siècles
suivants. Il existe d'importantes populations
sémitiques Hg J autour de la Méditerranée, mais
I est de plus grande incidence dans toute
l'Europe. La Grèce dispose également de la plus
grande population de pourcentage E3b en dehors
de l'Afrique.
Si le Haplogroupe I est vraiment sémite tel
qu'il apparaît alors les Bulgares et les
Roumains ont le plus grand pourcentage de
Sémites en Europe. Cependant, l'Europe du
Nord-Ouest et la Scandinavie ne sont pas loin
derrière eux. Les Anglo-Saxons ont apporté avec
eux quand ils sont venus du Moyen-Orient un
système de calendrier basé sur le sémite et il a
été appelé Al manach ou "le comptage" -
et notre mot Almanach
est dérivé de ce système. Un
Almanach original bimensuel de 59 jours basé sur
les Nouvelles Lunes est conservé à Cambridge.
Il semble que la Horde parthe était un composite
de la horde celtique et sémite. Ils étaient
devenus un peuple de petites tribus qui se sont
déplacées et recombinées pour s'installer dans
les régions de l'Europe. Leur langue était
teutonique et ils ont absorbé les Celtes qui les
avaient précédés, mais ils étaient eux-mêmes un
grand pourcentage de Celtes venant des alliances
hittites qu'ils avaient formées au Moyen-Orient.
Penchons-nous maintenant sur les divisions qui
semblent provenir de l'Haplogroupe K à partir
duquel les tribus japhétiques semblent être
descendues.
Les Fils de Japhet
Les Haplogroupes K, L, M, N, O, P, Q et R
semblent être les tribus japhétiques. Dieu dit
qu'Il bénirait Japhet. Il a été dit que Dieu
l’élargirait et qu'il habiterait dans les tentes
de Sem. Ainsi, on pourrait s'attendre à trouver
un mélange de Japhet et de Sem dans les mêmes
groupes nationaux. Ainsi les regroupements des
Hgs I et R1b et R1a répondent à ces critères.
K est la division racine de toutes les divisions
ultérieures de l’Haplogroupe de L à R2.
La majorité de K est en
Papouasie-Nouvelle-Guinée et en Mélanésie et
dans les Chinois Han. Il y a des petits
pourcentages parmi les Ouïgours, les Ouzbeks et
les Mongols et chez les Polynésiens. Certains K
se retrouvent également parmi les Italiens.
Ainsi cette division racine s’est déplacée
progressivement à l'est, avec une petite
quantité restant dans la Méditerranée, en Grèce
et en Italie. Les autres groupes ont muté de ce
Haplogroupe.
Ceux-ci sont les fils de Japhet : Gomer, et
Magog, Madaï, Javan, Tubal, Méschec et Tiras.
Les fils de Gomer : Ashkenaz, Riphath et
Togarma. Ce sont les Gimirra mentionnés
par les Assyriens qui étaient les Cimmériens
d'Hérodote. Ils sont les ancêtres des Celtes.
Les Ashkénazes ont formé la Horde Khazar. Les
Riphathiens ont formé les Celtes de l'Ouest ou
troyens de Wilusia. Ils sont devenus les anciens
Bretons à partir de 1000 avant notre ère, se
déplaçant ou subjuguant les Celtes Magogites en
Grande-Bretagne. Les Anglo-Saxons et les Jutes à
leur tour les ont déplacés à partir du cinquième
siècle de l'ère actuelle. Ils ont repoussé
l'invasion des Huns de Grande-Bretagne au 10ème
siècle, quand Humber roi des Huns a été tué
là-bas à la bataille de la rivière Humber nommée
d'après lui.
Magog est associé à Gomer dans Ézéchiel 38:2,6.
Magog est donc un ancêtre des Celtes aussi, et
ils sont les Milésiens qui ont envahi l'Irlande
plus tard dans le quatrième/cinquième siècle AEC
et les Celtes Scythes qui sont venus en Écosse,
au cinquième siècle EC. Josephus dit que Magog
est l'ancêtre des Scythes (A des J, Livre
1, ch. vi.). La distinction dans la langue
celtique de ces groupes est appelée brittonique
P et Q, elles ne sont pas la même langue.
Cependant, ils sont venus d'Anatolie et dans les
régions au nord dans ce qui était la Scythie.
L'alliance hittite a eu de nombreuses langues et
les Grecs décrivent les formations de l'armée de
Troie comme étant de nombreuses langues.
Nous savons également du récit historique au
Royaume-Uni que les Basques ont fait partie des
premiers habitants de la région ibérique de ce
qui est aujourd'hui le nord de l'Espagne et sont
prétendument descendants de Godel. Godel ou les
Godeliens sont les descendants de Japhet. Les
histoires disent aussi qu'ils sont les
descendants de Tubal et de Tarsis (Tarshish), le
fils et petit-fils de Japhet (Catholic
Encyclopaedia,
Vol. XIV, p. 176). Tarsis était le fils de
Javan, et non de Tubal. Ainsi, les histoires
soutiennent qu’il y avait deux groupes
japhétiques en Espagne. La Bible identifie les
Tarsis (2Chroniques 9:21 ; 20:36,37 ; Ps.
48:7 ; Ésaïe 2:16 ; 23:1,10,14 ; 60:9 ; Jér.
10:9 ; Ézéchiel 27:12,25 ; 38:13 ; Jonas 1:3 ;
4:2). Ce sont les Ibères, qui ont été divisés
dans les Basques dans le secteur nord et les
Tartesiens dans le sud (ibid.). Les Celtes se
sont ensuite superposés sur ces deux populations
antérieures ou autochtones. Ensuite, les Celtes
ont été dans le nord et l'ouest et les Ibères, à
l'est et au sud, avec la fusion des "Celtibères"
dans le centre d'Aragon et de Castille.
Cependant, ils ont aussi un élément de I parmi
eux, quoique plus faible que celui observé en
Irlande à environ >7%. Les Phéniciens ont
également établi des colonies là-bas sur les
côtes de Cadix, qui était leur capitale, et
aussi à Carteya, Calpe, Malaga et Sexi. Les
Rhodiens ont également établi une colonie à
Rosas au sud du cap Cresus ca. 910 avant notre
ère, et les Phocéens à Emporium. Les
Carthaginois s'installèrent dans les îles
Baléares au septième siècle avant notre ère. Au
sixième siècle, ils ont aidé les Phéniciens
contre les Tartesiens et ont ensuite pris
possession de la ville principale de Cadix et
ont commencé à faire du commerce en Beatica. Ils
ont commencé la conquête de l'Espagne après les
pertes de la Sicile dans la Première Guerre
Punique, mais ils l’ont perdue complètement
après la Deuxième Guerre Punique.
La langue basque est considérée comme être du
groupe linguistique ouralo-altaïque et de là est
reliée au finnois et à l'altaïque asiatique, au
mongol, au mandchou-Toungouses et au japonais.
Les Basques sont cependant R1b et quelques I.
Les commentaires ci-dessus concernant les Ibères
montrent que Tubal a fondé les Ibères, et
Josephus dit (ibid.) qu'ils étaient les fils de
Thobel
–
d'où Tubal et Thobel sont tenus d'être les
mêmes. À l'origine, les fils de Méschec étaient
en Cappadoce (ibid.). Les Ibères ultérieurs
étaient les Goths de l'Ouest (Wisigoths) qui
sont venus avec la horde au deuxième siècle EC.
Nous avons identifié les Mèdes comme les Kurdes
d'aujourd'hui, et Bullinger affirme que cela
doit être le cas. Il dit qu'ils étaient les
Amada ou Amadai des monuments assyriens (environ
840 avant notre ère), et donc maintenant les
tribus kurdes qui étaient à l’est de l'Assyrie.
Togarma est décrit comme étant dans les
parties les plus reculées du Nord et semble
être le Haplogroupe N et ses variations. Les
fils de Togarma, si cela devait être le cas,
serait la subdivision M214 de K suite à la
scission de N dans N (LLY22). Les Chinois et les
Malais sont O (M175). Le groupe ouralo-altaïque
et les Chinois Han seraient donc des fils de
Japhet avec un grand mélange de Mongols
cuschites des Haplogroupes C et D et leurs
femmes.
Les fils de Javan étaient : Elischa, Tarsis,
Kittim et Dodanim. La Bible dit alors que par
ceux-ci ont été peuplées les îles des nations
selon leurs terres, chacun selon sa langue,
selon leurs familles dans leurs nations (Genèse
20:2-5).
Ce sont ces groupes avec les Phéniciens E3b de
Tyr, de Sidon et de Carthage qui ont formé
l'empire commercial des Rois Anciens de la Mer.
Le sacerdoce des Crétois avait la peau
légèrement plus claire que les autres, comme
nous le voyons maintenant des reliefs du palais
de Cnossos, et ils semblent avoir aussi adoré le
Veau d'Or, d’après les boucles d'oreilles dans
le musée en Crète. Il y a un pourcentage
important de E3b autour des rives de la
Méditerranée.
Les fils de Javan ont été identifiés comme les
Ioniens de l'écriture cunéiforme et les tables
de Tel el-Armana. Les Grecs ont d'abord été
connus sous ce nom, mais les divisions d'ADN des
Grecs modernes indiquent que nous avons affaire
à un peuple différent des Grecs ioniens
d'origine.
Les fils d’Élischa sont compris comme étant les
Alasia des Égyptiens
–
en d'autres termes, les habitants de la côte de
Cilicie. Bullinger pense que Tarsis est
probablement Tarse. Cependant, Tarsis était en
Ibérie ou ce qui est aujourd'hui le sud de
l'Espagne. C'est à partir de ce port que la
plupart du commerce transatlantique a été
effectué dans les temps anciens. Les Kittim
étaient les ancêtres des anciens empires
commerciaux marins dirigés à partir de Chypre.
Les Dodanim sont compris comme étant les
Rhodanim (1Chroniques 1:7, cf. Septante).
Méschec et Tubal ont été associés les uns aux
autres. Les Tabali sont associés comme et avec
Tubal. Méschec est associé aux Muska des
monuments assyriens. Nous comprenons ceux-ci
comme les Russes et leurs Haplogroupes sont
principalement R1a avec une petite quantité de
R1b et de N, I et J significatifs, et certains
E3b dans le nord de la Russie.
Ces tribus étaient à l'origine dans le nord de
l'Asie Mineure au-dessus des Magogites, mais
avec la défaite des Hittites et la chute de
Troie, ils sont entrés en Anatolie en Cappadoce.
C'est le conflit évoqué par le Roi David quand
il est avec Méschec (et peut-être Tubal). Les
tribus étaient beaucoup plus au sud en
Géorgie/Arménie où il écrivait ca. 1000 avant
notre ère. La zone autour de la Géorgie a
également été appelée Iberia à partir
d’Iber. Cela peut être la source de ce nom, et
représente l'utilisation du nom en Espagne.
Nul doute que David a essayé de contenir Méschec
(et peut-être Tubal) comme il a mentionné que
quand il est pour la paix, ils sont pour la
guerre (Ps. 120:7). Il mentionne aussi les
tentes de Kédar. Ézéchiel 38:2 se réfère au chef
de Ro'sh, Méschec et de Tubal. Ro'sh signifie
également chef ou prééminent et
donc il est traduit dirigeant en chef ou
prince plutôt que comme un nom ; mais il
peut être à l'origine du terme Russ. Les
Vikings Russ de 850 à 950 EC ont formé le
Royaume Varangian des Russ autour de Kiev et de
Novgorod. Il y a des éléments R1a en Suède et en
Norvège et bien plus encore qu'ailleurs en
Europe occidentale, mais les Slaves sont les
vrais groupes R1a, avec le plus grand
pourcentage chez les Polonais. Les Ro'sh ont été
associés avec la Russie, et les villes de Muskva
(Moscou) et de Tobolsk ont été associées avec
les tribus. Ils se sont déplacés vers le nord
après leur confinement au début du premier
millénaire avant notre ère. Néanmoins, ils ont
aussi une importante population de l’Haplogroupe
I. Quand Ézéchiel se réfère à Gog et le prince
Ro'sh de Méschec et de Tubal il dit qu'ils sont
dans les pays de Magog. Autrefois cette région
se trouvait au nord de l'Anatolie et à l'est de
la zone de Troie dans ce qui est maintenant la
Turquie, l'Arménie et la Géorgie.
Tiras était l'habitant d'origine de la Thrace (à
partir de Tirasia), mais la tribu de ses
descendants ne semble pas y être restée puisque
les Haplogroupes semblent être sémites.
Il y a un Haplogroupe qui s'est développé à
partir de l'Haplogroupe P (M45, M74, P27, 92R7,
P69?) : qui est Q (M242, MEH2, P36). C'est à
côté du supergroupe de l’Haplogroupe R. Q se
produit au sein des Selkups et de l'Altaï en
Russie et va ensuite directement dans la Sibérie
et l'océan Pacifique en Amérique. Il existe une
autre façon par laquelle la tribu aurait pu se
déplacer et cela est par la mer de l'autre côté
de l'Atlantique.
Nous savons pour un fait, maintenant que
l'expédition transatlantique se passait sur une
base régulière ca. 1000 avant notre ère entre
l'Égypte et l'Amérique. De récents tests
médico-légaux sur les momies du musée de Berlin
se sont avérés positifs à la cocaïne et la
consommation de tabac sur chaque momie au musée.
Le British Museum a soupçonné que les résultats
étaient contaminés et a testé l'ensemble de
leurs momies pour la comparaison et ils n'ont
pas trouvé la cocaïne dans les leurs, mais
chacune était positive pour la consommation de
tabac. Or, ces produits ne poussent que dans
l'Amérique. Les momies étaient toutes autour de
1000 avant notre ère environ. Le commerce semble
avoir continué au cours du deuxième millénaire
avant notre ère jusqu'au premier millénaire EC,
comme Cyrus Gordon montre à partir de la preuve.
Le commerce était dans le Pacifique et
l'Atlantique.
Peut-être que les Amérindiens se sont déplacés
comme décrit ci-dessus, mais ils faisaient
certainement du commerce à travers l'Atlantique
avec les Tarsis ibériques et les Rois Anciens de
la Mer à l'époque du Roi David, et jusqu'à
l’époque des Romains. Il existe des preuves de
commerce romain là au premier siècle de l'ère
actuelle. Les rebelles Bar Kochba seraient
également allés là-bas au deuxième siècle de
l'ère actuelle et leurs pièces de monnaie et
leurs tombes ont été retrouvées dans le Midwest
des États-Unis (voir également le professeur
Cyrus Gordon, Before Columbus,
Touchstone, 1972).
Une des choses bizarres sur les traversées
transatlantiques, c'est qu'il y a des tribus qui
sembleraient avoir des liens avec les
Britanniques gallois ou antiques, mais nos
dossiers actuels ne montrent pas le Haplogroupe
R1b en quantités. Ces aspects doivent être
étudiés davantage. L'ADN des différentes tribus
indiennes et en particulier celles de la
Virginie et le nord-est a besoin d’être
quantifié, comme le script Ogham aurait été
trouvé là - et si l'ADN devait révéler un
contact s’il a eu lieu. Les tests d’ADN-Y
effectués sur les Indiens d'Amérique du Nord à
ce jour ont révélé Q, Q3 et C3 uniquement.
Comme nous l'avons déjà vu ; le Haplogroupe R a
RxR1 de base de l'Afrique à l'Inde à
l'Australie.
R1a comprend la majorité des Slaves et des
Indo/Aryens tandis que le Haplogroupe R1b
comprend la majeure partie des Européens de
l'Ouest, à la fois Celtes et
Anglo-Saxons/Normands. R1a s'étend en Allemagne
et en Norvège en plus grande quantité que trouvé
nulle part ailleurs en Europe occidentale. En
Islande, nous voyons R1a, R1b et I commencer à
fusionner avec Q mais seulement ici (à partir
d’infusion Eskimo là-bas et à certaines familles
en Norvège).
Si le Haplogroupe I est le Haplogroupe définitif
sémitique, alors les Sémites sont répartis à
travers l'Europe, mélangés avec Japhet. Les
Basques, les Gallois et les Irlandais sont les
peuples les plus distincts avec un Haplogroupe
simple (R1b), mais même là ils ont jusqu'à
quatre haplogroupes distincts, avec les Basques
et les Gallois/Cornouaillais ayant seulement
deux Haplogroupes.
L'argument pourrait être soulevé que les
Haplogroupes de Sem et Japhet sont à la fois F
et que tous leurs fils sont de simples
variations d’eux, sans distinction, et donc G à
J sont de deux groupes, avec K formant la
division suivante et qui n’apparaît pas dans le
célèbre J sémite. Ainsi K doit inclure tout
Japhet, mais il pourrait être alors fait valoir
que G à I est ouvert aux deux. Ces arguments ne
semblent pas tenir un examen sur une base
démographique simple.
Les Évolutionnistes affirment que ces lignées
sont toutes sorties en rampant des marais avec
des ancêtres en vie jusqu'à il y a 130.000 ans.
Ainsi, J est la lignée sémite et les autres sont
toutes les activités indépendantes de
l'évolution, mais ils avouent que tout cela
vient d'un seul ancêtre. Ils soutiennent que le
Déluge est un mythe et que toute personne qui
prend au sérieux le récit biblique est tout
simplement ignorante de la réalité scientifique.
Pour que la Bible soit la Parole inspirée de
Dieu, elle doit expliquer l'origine fondamentale
des nations d'une manière rationnelle et vraie.
Ce que nous avons vu ici c’est que l'ADN de
groupes tribaux connus correspond à l'ADN
d'autres groupes qui sont identifiés avec leurs
proches bibliques.
L'autre argument est que tout arrive simplement
beaucoup plus vite que nous le pensions et que
ces variations se sont produites indépendamment
dans plusieurs endroits, et la séquence d'ADN
n'est pas une indication de l'origine des
personnes. À court terme, cette hypothèse n'est
pas en accord avec le test connu et les exemples
historiques.
Ce qui a été proposé ici est en accord avec
l'histoire connue et le récit biblique. Cela
fournit une explication en utilisant la
génétique qui est explicable dans le récit
biblique et est une alternative acceptable aux
modèles évolutifs utilisant des périodes de
temps étirées.
Il est également possible que le Haplogroupe I
soit le Haplogroupe majeur sémitique couvrant
les Hébreux et s'étendant vers les Israélites du
Nord et aussi aux Élamites. Alors que nous
n'avons trouvé aucun Haplogroupe I chez les
Assyriens modernes, plutôt que G et J avec L,
certains de l’Hg I ont pu se produire autrefois.
La prophétie selon laquelle Israël et l'Assyrie
sortiront main dans la main du septentrion dans
les Derniers Jours à la restauration peut se
produire parce qu'ils sont tous deux dispersés
en Europe et certains peuvent même être Hg I ;
mais les tests modernes ne montrent aucun Hg I
dans ces groupes identifiés comme Assyriens
modernes. Le Haplogroupe J est donc un
développement ultérieur du Moyen-Orient dans les
pays arabes après la captivité babylonienne et
couvre les Juifs, les Ismaélites et les Édomites
absorbés dans Juda. Même sur les modèles
évolutionnistes, qui sont très biaisés aux plus
anciennes dates possibles AEC, le Haplogroupe
Modal Cohen (CMH) J2 est admis pour être vieux
d’un 2100 ans possible. Ainsi l’Hg I pourrait
très bien l’avoir précédé de plusieurs siècles
et identifier les Dix Tribus d'Israël, qui sont
ensuite réparties dans toute l'Europe. La
bénédiction de Japhet place ses descendants dans
les tentes de Sem et donc la prophétie est
accomplie et les bénédictions de Sem reviennent
à Japhet, même si Japhet est plus grand en
termes de pourcentage dans chaque nation.
Ce qui est certain, c'est que dans la mesure
même des Haplogroupes I et R1b et R1a aussi en
Europe, il est pratiquement impossible pour
presque chaque personne en Europe de ne pas être
un descendant d'Abraham, d'Isaac et de Jacob,
que ce soit à travers leurs lignées maternelles
ou paternelles. Ainsi l'alliance hittite avec
Israël en a fait un seul peuple réparti sur une
vaste zone avec des entités linguistiques.
Abraham est en effet devenu le père de beaucoup
de nations.
Lisez également les documents suivants :
Les Fils de Japhet : Partie I
(No. 46A)
Les Fils
de Japhet Partie 1A : Les Fils de HN (No. 046A1)
Les Fils
de Japhet Partie II : Gomer (No. 046B)
Les Fils
de Japhet Partie III : Magog (No. 046C)
Les Fils
de Japhet Partie V : Javan (No. 046E)
Les Fils
de Japhet : Partie VI Tubal (No. 046F)
Les Fils de
Japhet Partie VIII Tiras (No. 046H)
Les Fils de Cham : Partie I
Introduction (No. 45A)
Les Fils de Cham : Partie II Cush
(No. 45B)
Les Fils de Cham : Partie III
Mitsraïm (No. 45C)
Les Fils de Cham : Partie IV Phut
(No. 45D)
Les Fils de Cham : Partie V Canaan
(No. 45E)
Les Fils de Sem : Partie I
(No. 212A)
Les Descendants
d'Abraham Partie II : Lot, Moab, Ammon et Ésaü (No. 212B)
Les Descendants
d'Abraham Partie III : Ismaël (No. 212C)
Les Descendants
d'Abraham Partie IV : Les Fils de Ketura (No. 212D)
Les Descendants
d'Abraham Partie V : Juda (No. 212E)
Les Descendants
d'Abraham Partie VI : Israël (No. 212F)
Les Fils de Sem : Partie VII
: Tableaux pour 212A-212F (No. 212G)
Les Descendants d'Abraham
Partie VIII : Les Treize Famines de la Rébellion (No. 212H)
q